

Complement inhibition by Sarcoptes scabiei protects Streptococcus pyogenes - An in vitro study to unravel the molecular mechanisms behind the poorly understood predilection of S. pyogenes to infect mite-induced skin lesions
- PMID: 28278252
- PMCID: PMC5360341
- DOI: 10.1371/journal.pntd.0005437
Complement inhibition by Sarcoptes scabiei protects Streptococcus pyogenes - An in vitro study to unravel the molecular mechanisms behind the poorly understood predilection of S. pyogenes to infect mite-induced skin lesions
Abstract
Background: On a global scale scabies is one of the most common dermatological conditions, imposing a considerable economic burden on individuals, communities and health systems. There is substantial epidemiological evidence that in tropical regions scabies is often causing pyoderma and subsequently serious illness due to invasion by opportunistic bacteria. The health burden due to complicated scabies causing cellulitis, bacteraemia and sepsis, heart and kidney diseases in resource-poor communities is extreme. Co-infections of group A streptococcus (GAS) and scabies mites is a common phenomenon in the tropics. Both pathogens produce multiple complement inhibitors to overcome the host innate defence. We investigated the relative role of classical (CP), lectin (LP) and alternative pathways (AP) towards a pyodermic GAS isolate 88/30 in the presence of a scabies mite complement inhibitor, SMSB4.
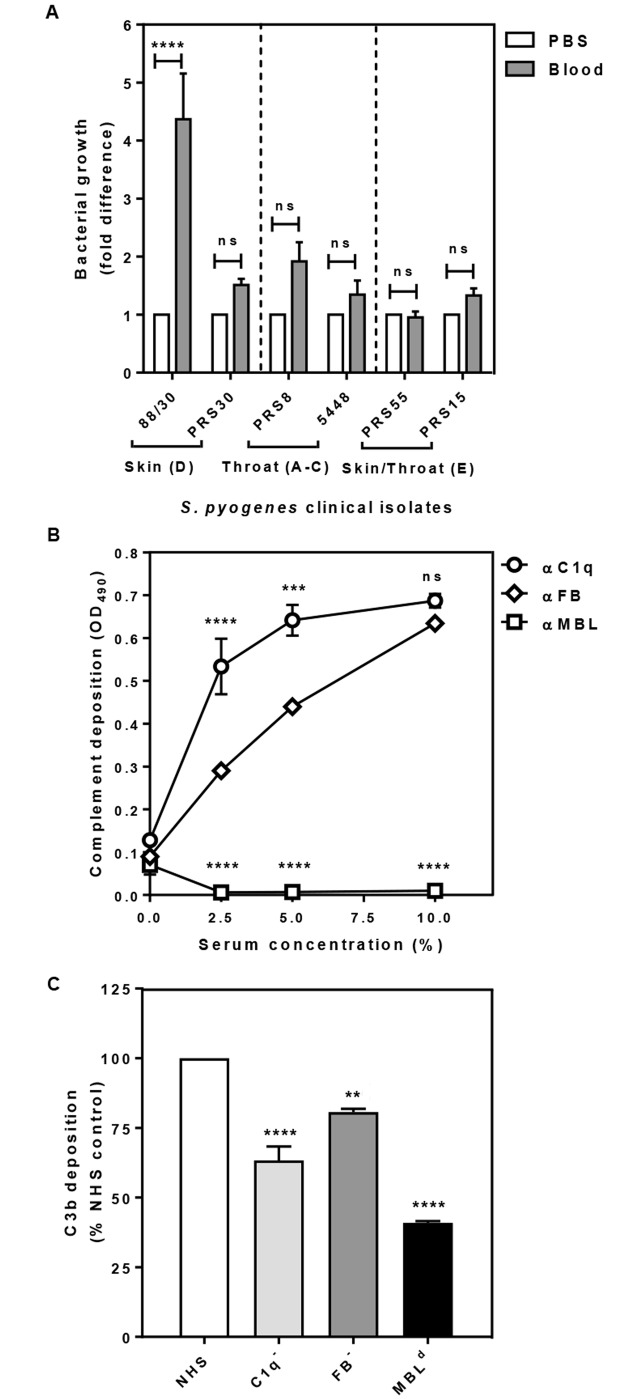
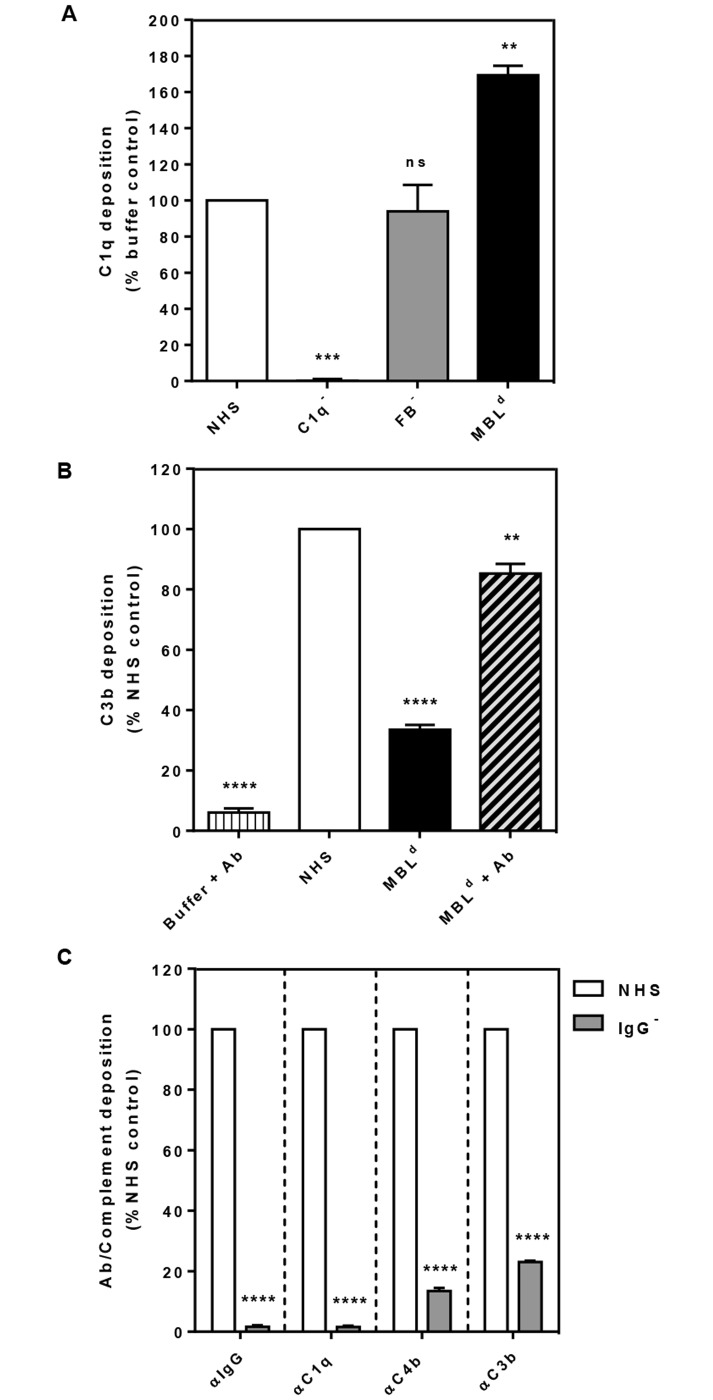
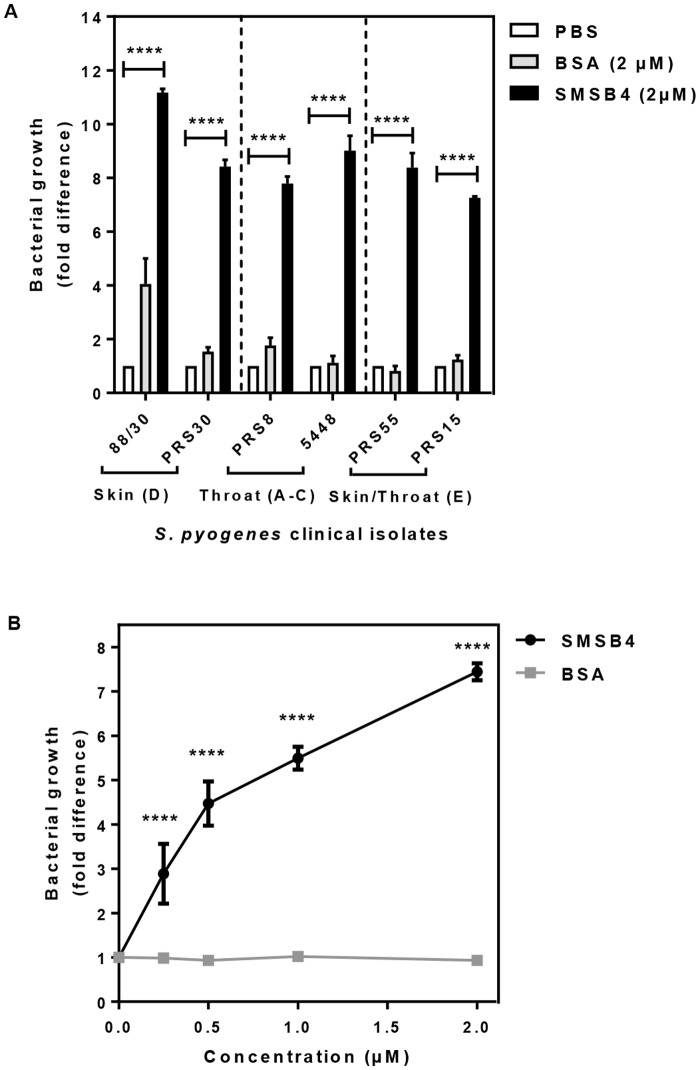
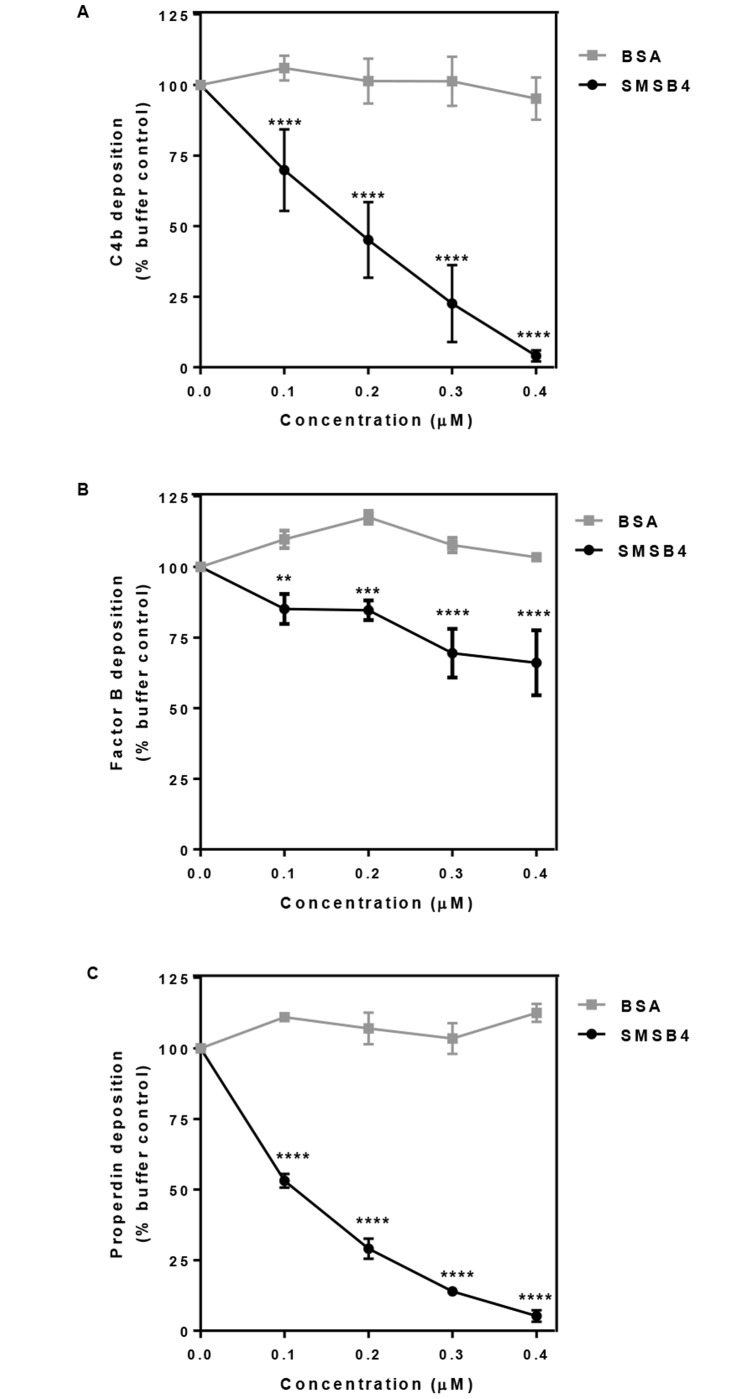
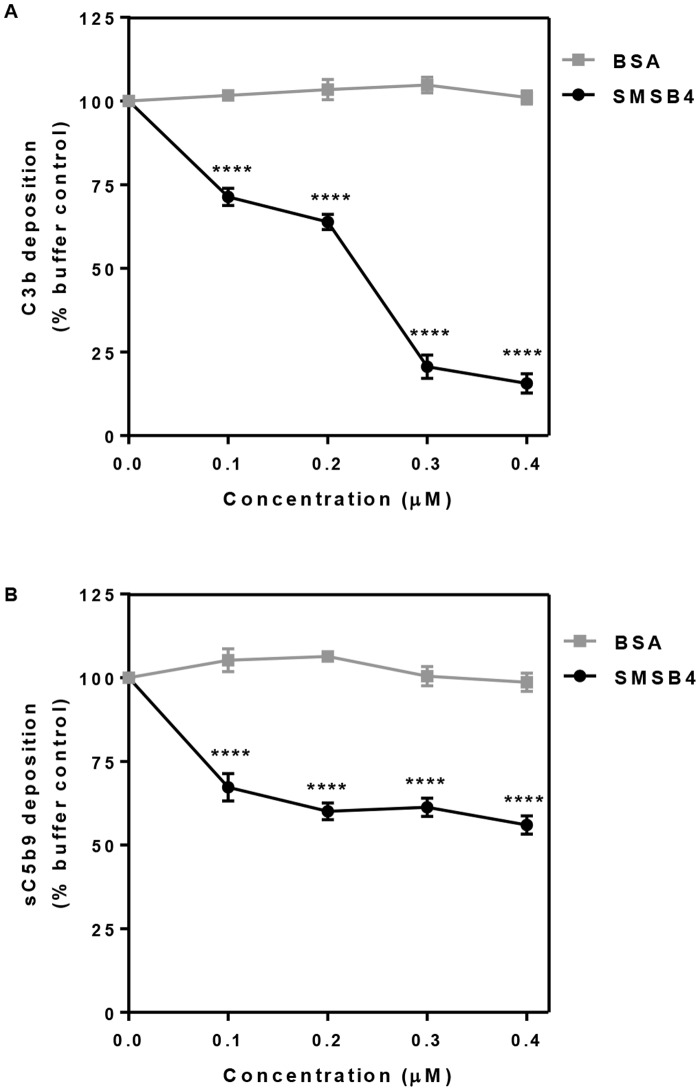
Methodology/principal findings: Opsonophagocytosis assays in fresh blood showed baseline immunity towards GAS. The role of innate immunity was investigated by deposition of the first complement components of each pathway, specifically C1q, FB and MBL from normal human serum on GAS. C1q deposition was the highest followed by FB deposition while MBL deposition was undetectable, suggesting that CP and AP may be mainly activated by GAS. We confirmed this result using sera depleted of either C1q or FB, and serum deficient in MBL. Recombinant SMSB4 was produced and purified from Pichia pastoris. SMSB4 reduced the baseline immunity against GAS by decreasing the formation of CP- and AP-C3 convertases, subsequently affecting opsonisation and the release of anaphylatoxin.
Conclusions/significance: Our results indicate that the complement-inhibitory function of SMSB4 promotes the survival of GAS in vitro and inferably in the microenvironment of the mite-infested skin. Understanding the tripartite interactions between host, parasite and microbial pathogens at a molecular level may serve as a basis to develop improved intervention strategies targeting scabies and associated bacterial infections.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Carapetis JR, Connors C, Yarmirr D, Krause V, Currie BJ. Success of a scabies control program in an Australian aboriginal community. Pediatr Infect Dis J. 1997;16(5): 494–9. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
