

Toll-Like Receptor 2 Is Required for Inflammatory Process Development during Leishmania infantum Infection
- PMID: 28280488
- PMCID: PMC5322192
- DOI: 10.3389/fmicb.2017.00262
Toll-Like Receptor 2 Is Required for Inflammatory Process Development during Leishmania infantum Infection
Abstract
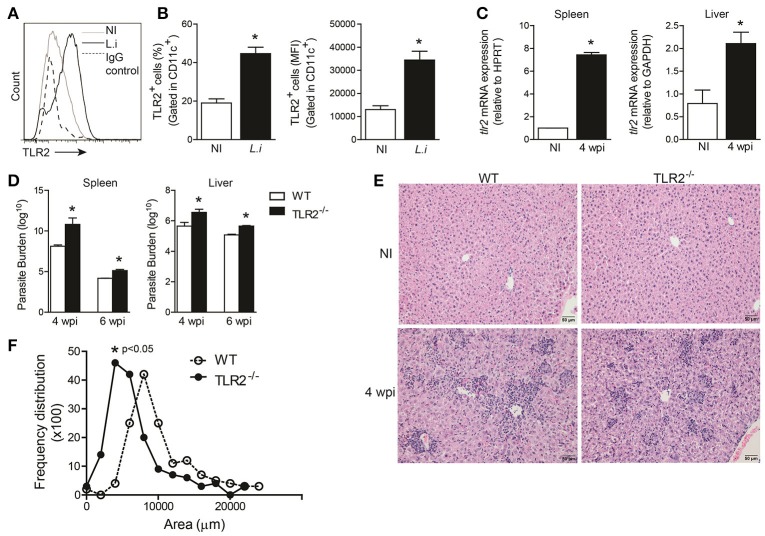
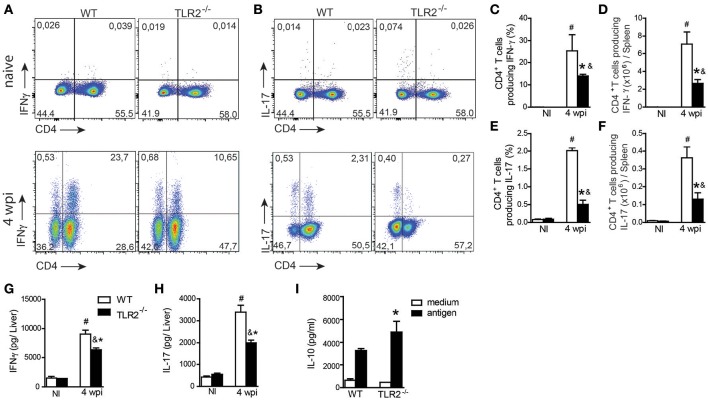
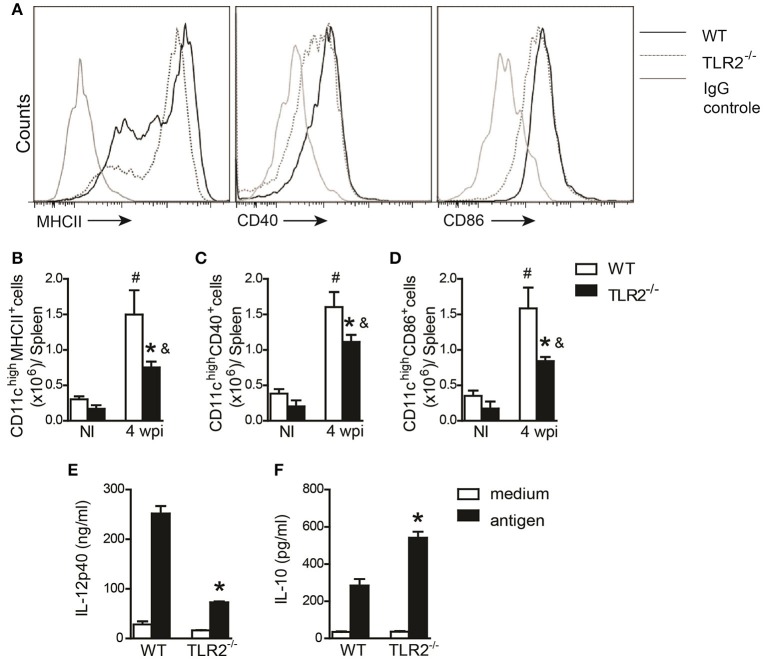
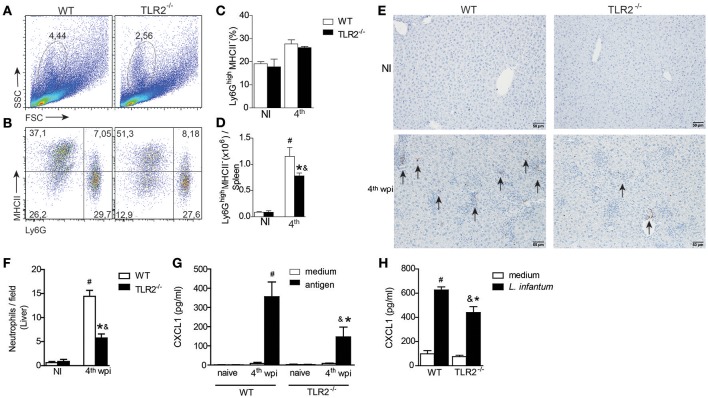
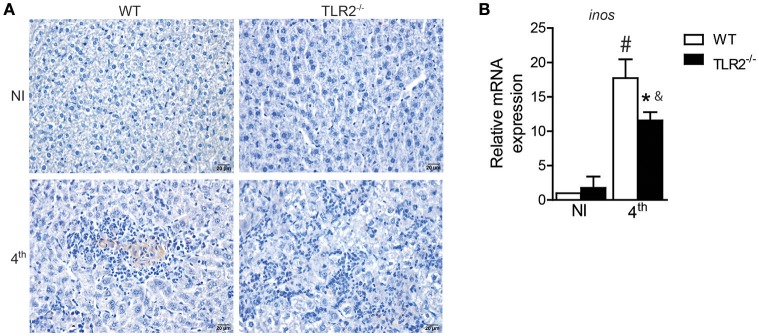
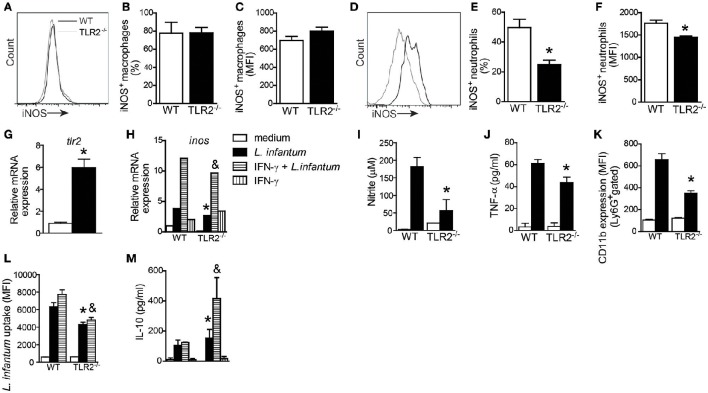
Visceral leishmaniasis (VL) is a chronic and fatal disease caused by Leishmania infantum in Brazil. Leukocyte recruitment to infected tissue is a crucial event for the control of infections such as VL. Among inflammatory cells, neutrophils are recruited to the site of Leishmania infection, and these cells may control parasite replication through oxidative or non-oxidative mechanisms. The recruitment, activation and functions of the neutrophils are coordinated by pro-inflammatory cytokines and chemokines during recognition of the parasite by pattern recognition receptors (PRRs). Here, we demonstrated that the Toll-like receptor 2 (TLR2) signaling pathway contributes to the development of the innate immune response during L. infantum infection. The protective mechanism is related to the appropriate recruitment of neutrophils to the inflammatory site. Neutrophil migration is coordinated by DCs that produce CXCL1 and provide a prototypal Th1 and Th17 environment when activated via TLR2. Furthermore, infected TLR2-/- mice failed to induce nitric oxide synthase (iNOS) expression in neutrophils but not in macrophages. In vitro, infected TLR2-/- neutrophils presented deficient iNOS expression, nitric oxide (NO) and TNF-α production, decreased expression of CD11b and reduced L. infantum uptake capacity. The non-responsive state of neutrophils is associated with increased amounts of IL-10. Taken together, these data clarify new mechanisms by which TLR2 functions in promoting the development of the adaptive immune response and effector mechanisms of neutrophils during L. infantum infection.
Keywords: Leishmania infantum; TLR2; dendritic cell maturation; neutrophil activation; visceral leishmaniasis.
Figures
References
-
- Assis R. R., Ibraim I. C., Noronha F. S., Turco S. J., Soares R. P. (2012). Glycoinositolphospholipids from Leishmania braziliensis and L. infantum: modulation of innate immune system and variations in carbohydrate structure. PLoS Negl. Trop. Dis. 6:e1543. 10.1371/journal.pntd.0001543 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials