

A full lifespan model of vertebrate lens growth
- PMID: 28280571
- PMCID: PMC5319337
- DOI: 10.1098/rsos.160695
A full lifespan model of vertebrate lens growth
Abstract
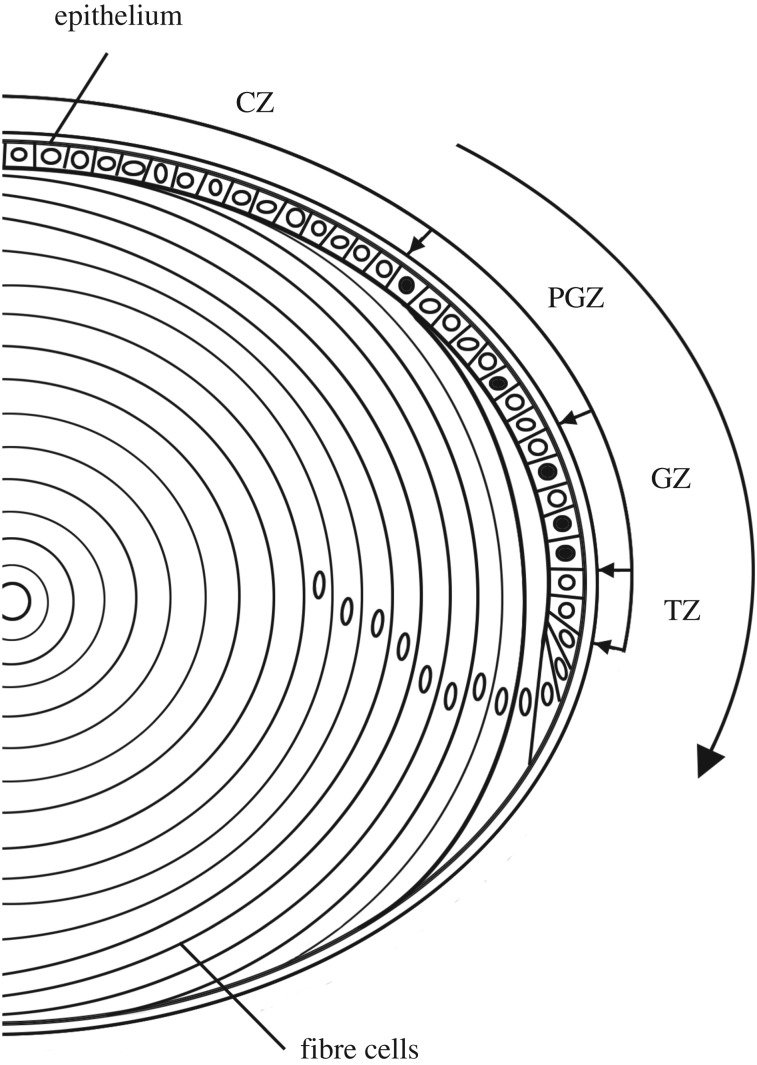
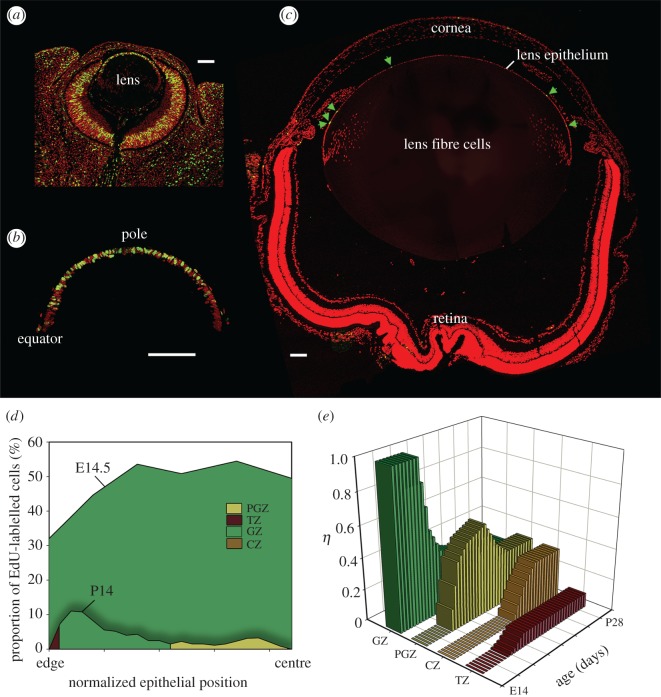
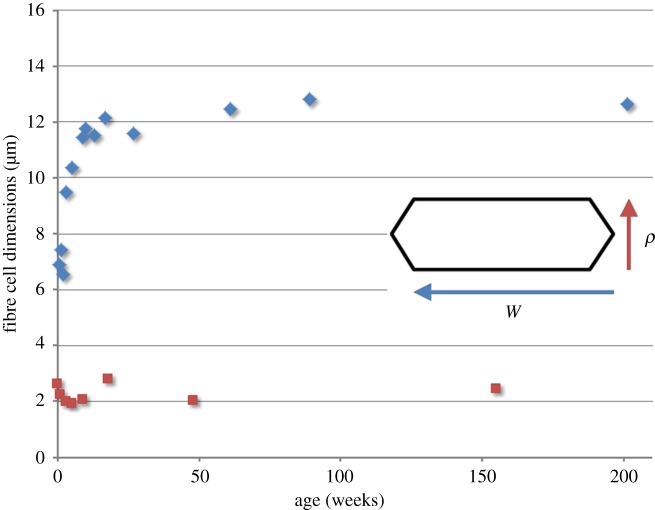
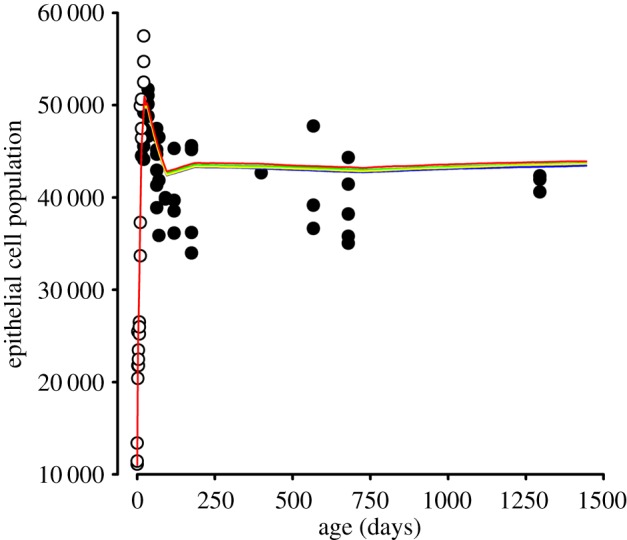
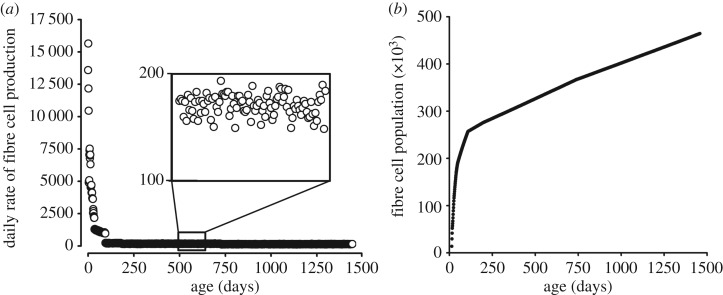
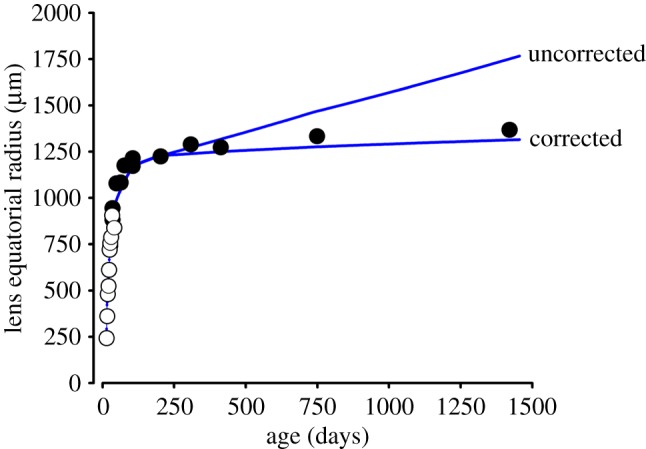
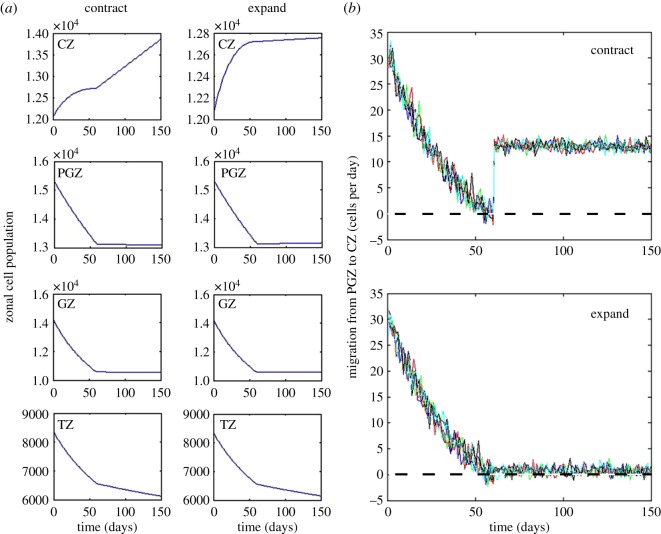
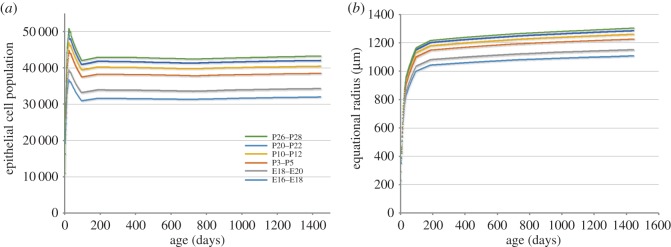
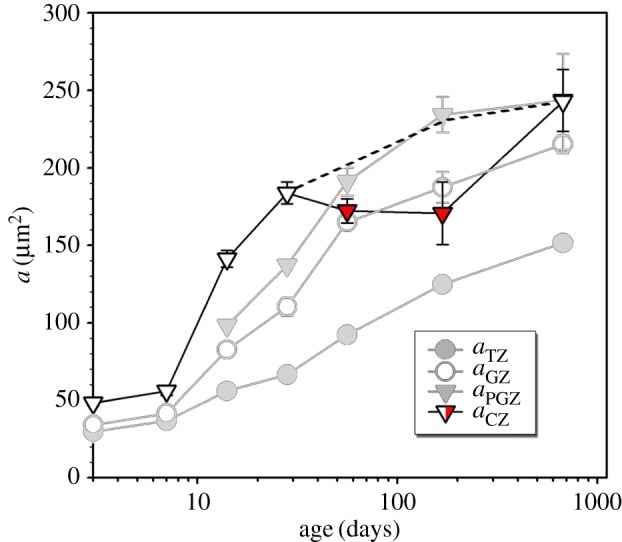
The mathematical determinants of vertebrate organ growth have yet to be elucidated fully. Here, we utilized empirical measurements and a dynamic branching process-based model to examine the growth of a simple organ system, the mouse lens, from E14.5 until the end of life. Our stochastic model used difference equations to model immigration and emigration between zones of the lens epithelium and included some deterministic elements, such as cellular footprint area. We found that the epithelial cell cycle was shortened significantly in the embryo, facilitating the rapid growth that marks early lens development. As development progressed, epithelial cell division becomes non-uniform and four zones, each with a characteristic proliferation rate, could be discerned. Adjustment of two model parameters, proliferation rate and rate of change in cellular footprint area, was sufficient to specify all growth trajectories. Modelling suggested that the direction of cellular migration across zonal boundaries was sensitive to footprint area, a phenomenon that may isolate specific cell populations. Model runs consisted of more than 1000 iterations, in each of which the stochastic behaviour of thousands of cells was followed. Nevertheless, sequential runs were almost superimposable. This remarkable degree of precision was attributed, in part, to the presence of non-mitotic flanking regions, which constituted a path by which epithelial cells could escape the growth process. Spatial modelling suggested that clonal clusters of about 50 cells are produced during migration and that transit times lengthen significantly at later stages, findings with implications for the formation of certain types of cataract.
Keywords: branching process; lens; model; organ growth; stochastic.
Figures
References
-
- Hariharan IK. 2015. Organ size control: lessons from Drosophila. Dev. Cell 34, 255–265. (doi:10.1016/j.devcel.2015.07.012) - DOI - PMC - PubMed
-
- Buchmann A, Alber M, Zartman JJ. 2014. Sizing it up: the mechanical feedback hypothesis of organ growth regulation. Semin. Cell Dev. Biol. 35, 73–81. (doi:10.1016/j.semcdb.2014.06.018) - DOI - PMC - PubMed
-
- Shi Y, De Maria A, Lubura S, Sikic H, Bassnett S. 2015. The penny pusher: a cellular model of lens growth. Invest. Ophthalmol. Vis. Sci. 56, 799–809. (doi:10.1167/iovs.14-16028) - DOI - PMC - PubMed
-
- Sikic H, Shi Y, Lubura S, Bassnett S. 2015. A stochastic model of eye lens growth. J. Theor. Biol. 376, 15–31. (doi:10.1016/j.jtbi.2015.03.021) - DOI - PMC - PubMed
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials