

Adipocytes promote malignant growth of breast tumours with monocarboxylate transporter 2 expression via β-hydroxybutyrate
- PMID: 28281525
- PMCID: PMC5353665
- DOI: 10.1038/ncomms14706
Adipocytes promote malignant growth of breast tumours with monocarboxylate transporter 2 expression via β-hydroxybutyrate
Abstract
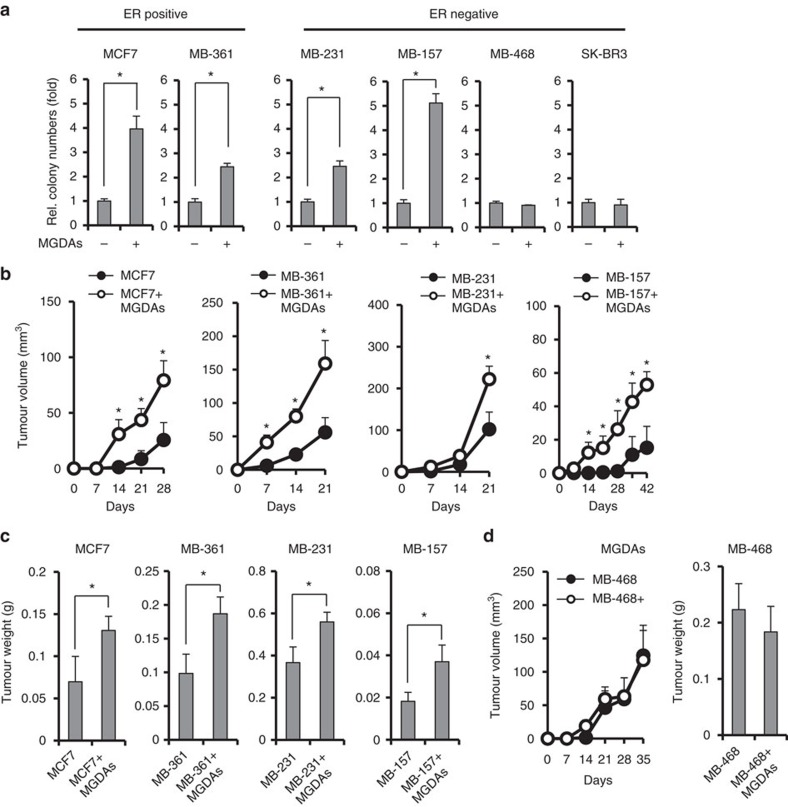
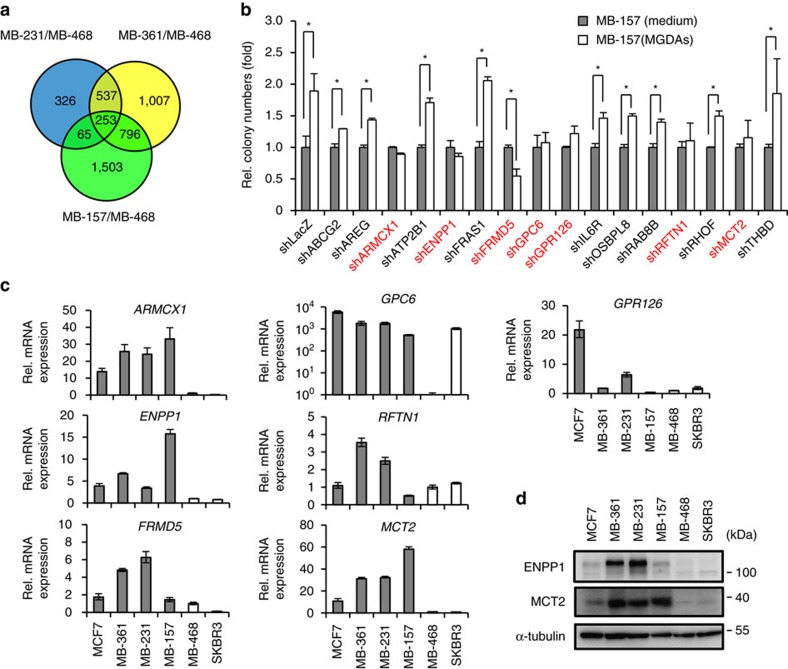
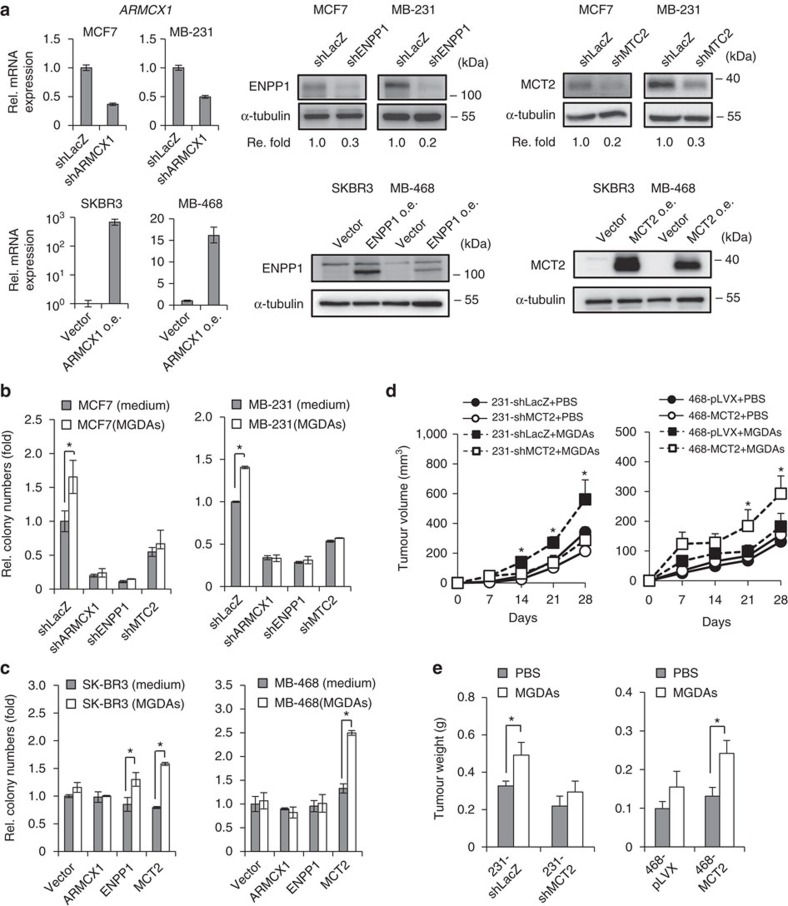
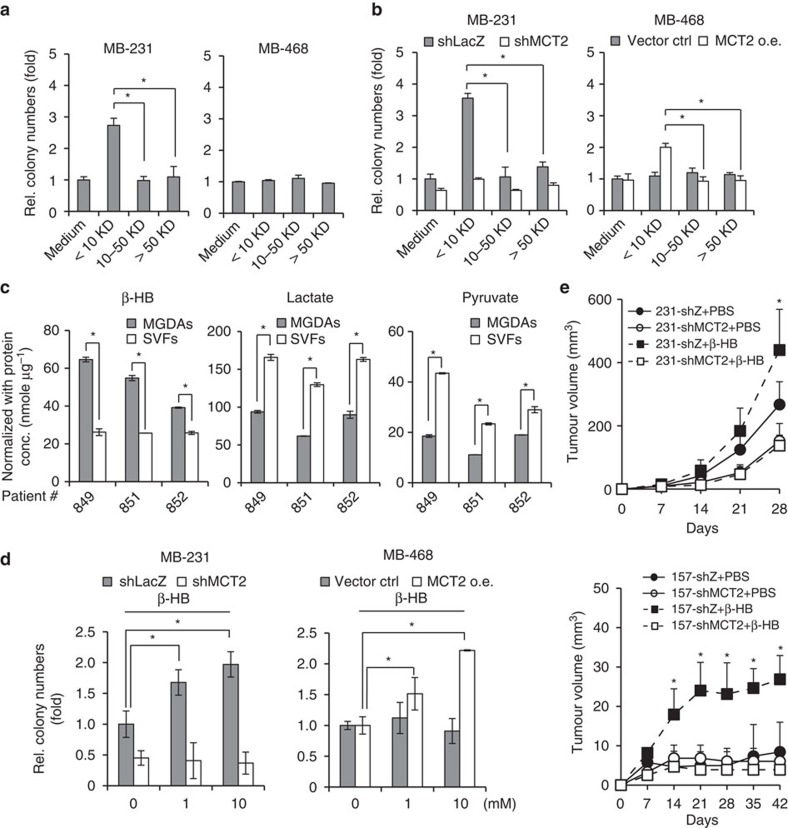
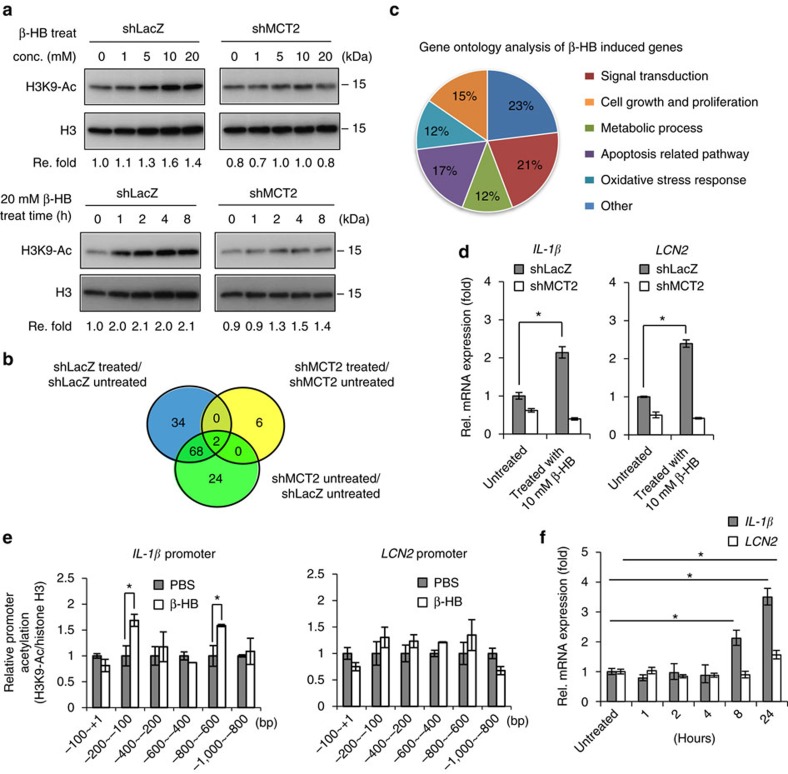
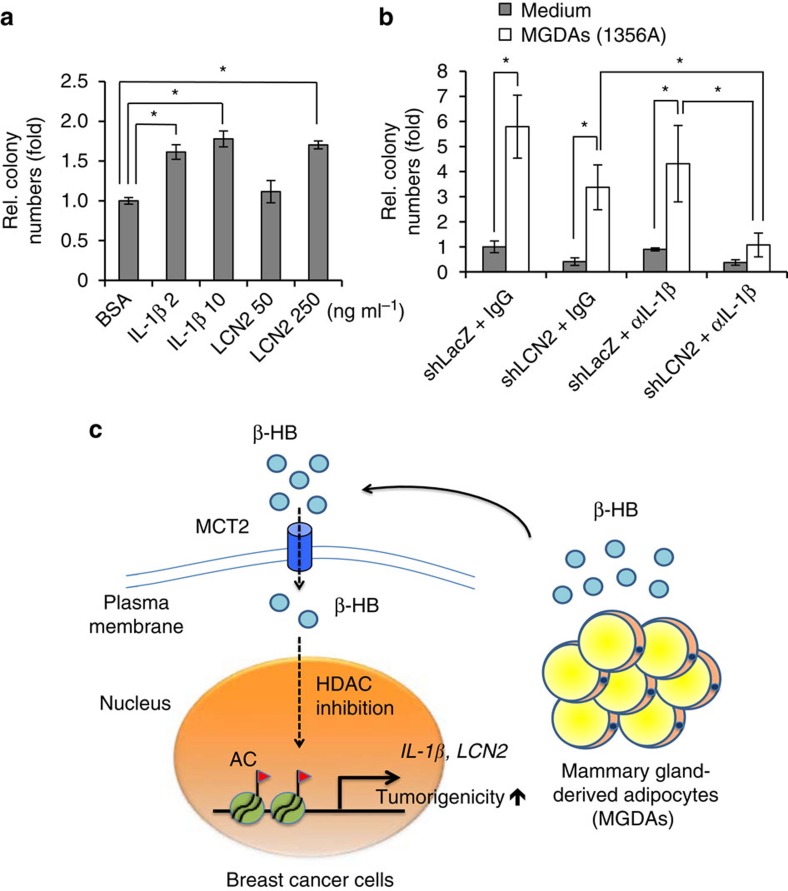
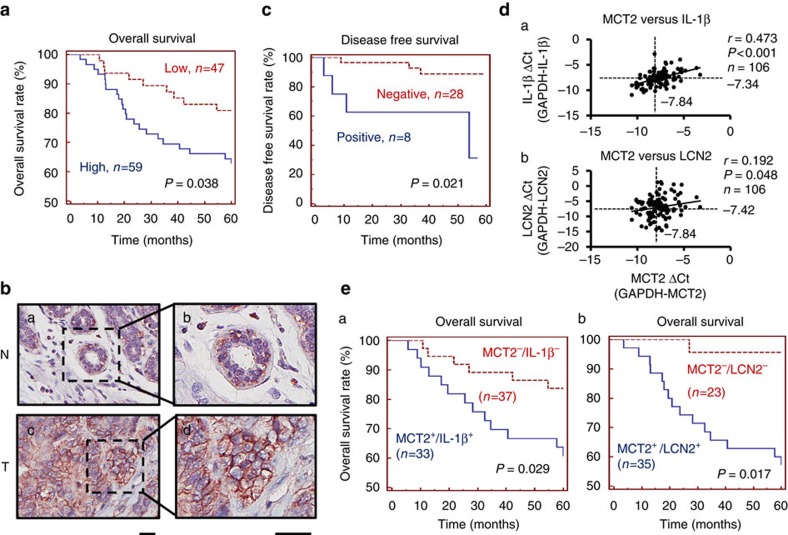
Adipocytes are the most abundant stromal partners in breast tissue. However, the crosstalk between breast cancer cells and adipocytes has been given less attention compared to cancer-associated fibroblasts. Here we find, through systematic screening, that primary mammary gland-derived adipocytes (MGDAs) promote growth of breast cancer cells that express monocarboxylate transporter 2 (MCT2) both in vitro and in vivo. We show that β-hydroxybutyrate is secreted by MGDAs and is required to enhance breast cancer cells malignancy in vitro. Consistently, β-hydroxybutyrate is sufficient to promote tumorigenesis of a mouse xenograft model of MCT2-expressing breast cancer cells. Mechanistically we observe that upon co-culturing with MGDAs or treatment with β-hydroxybutyrate, breast cancer cells expressing MCT2 increase the global histone H3K9 acetylation and upregulate several tumour-promoting genes. These results suggest that adipocytes promote malignancy of MCT2-expressing breast cancer via β-hydroxybutyrate potentially by inducing the epigenetic upregulation of tumour-promoting genes.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Liotta L. A. & Kohn E. C. The microenvironment of the tumour-host interface. Nature 411, 375–379 (2001). - PubMed
-
- Orimo A. et al.. Stromal fibroblasts present in invasive human breast carcinomas promote tumor growth and angiogenesis through elevated SDF-1/CXCL12 secretion. Cell 121, 335–348 (2005). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
