

Extensive translation of circular RNAs driven by N6-methyladenosine
- PMID: 28281539
- PMCID: PMC5520850
- DOI: 10.1038/cr.2017.31
Extensive translation of circular RNAs driven by N6-methyladenosine
Abstract
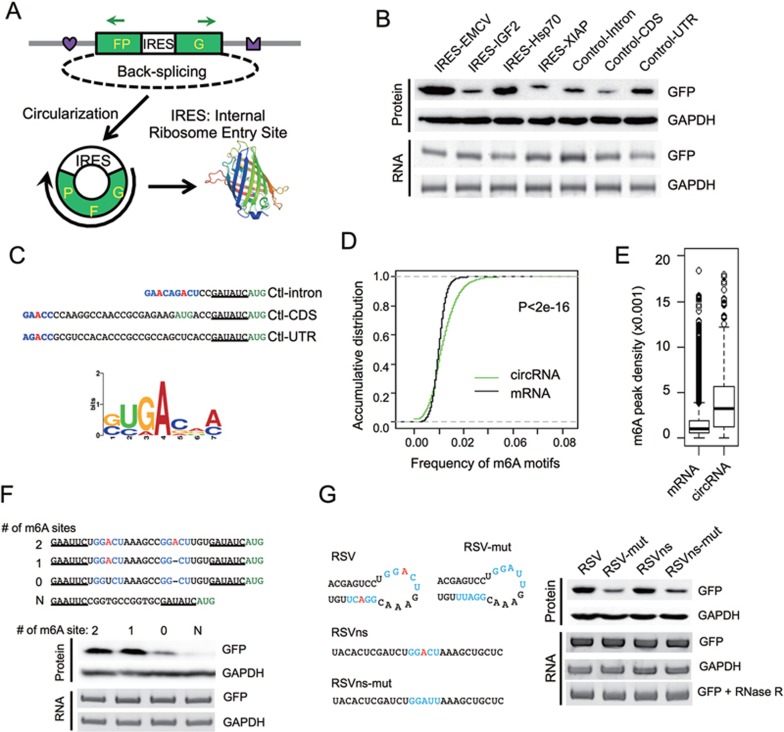
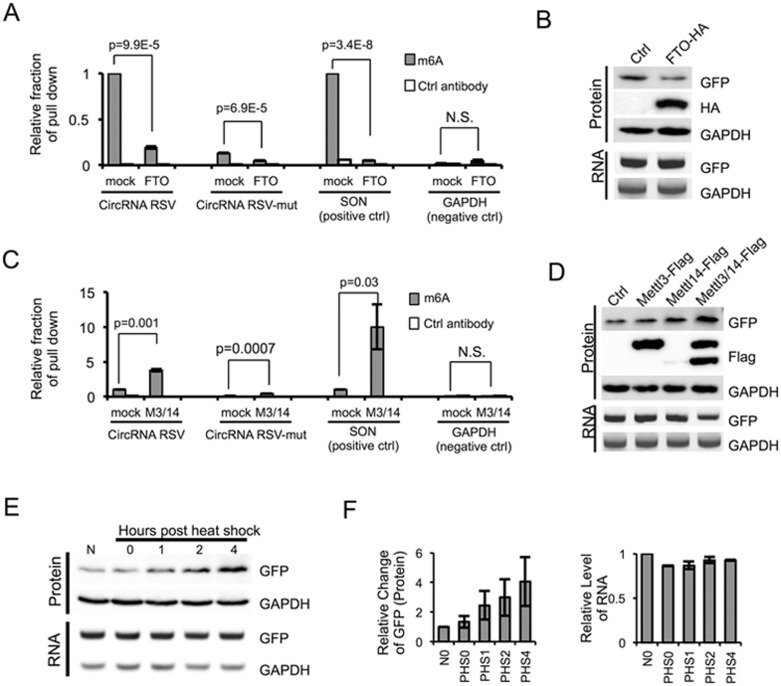
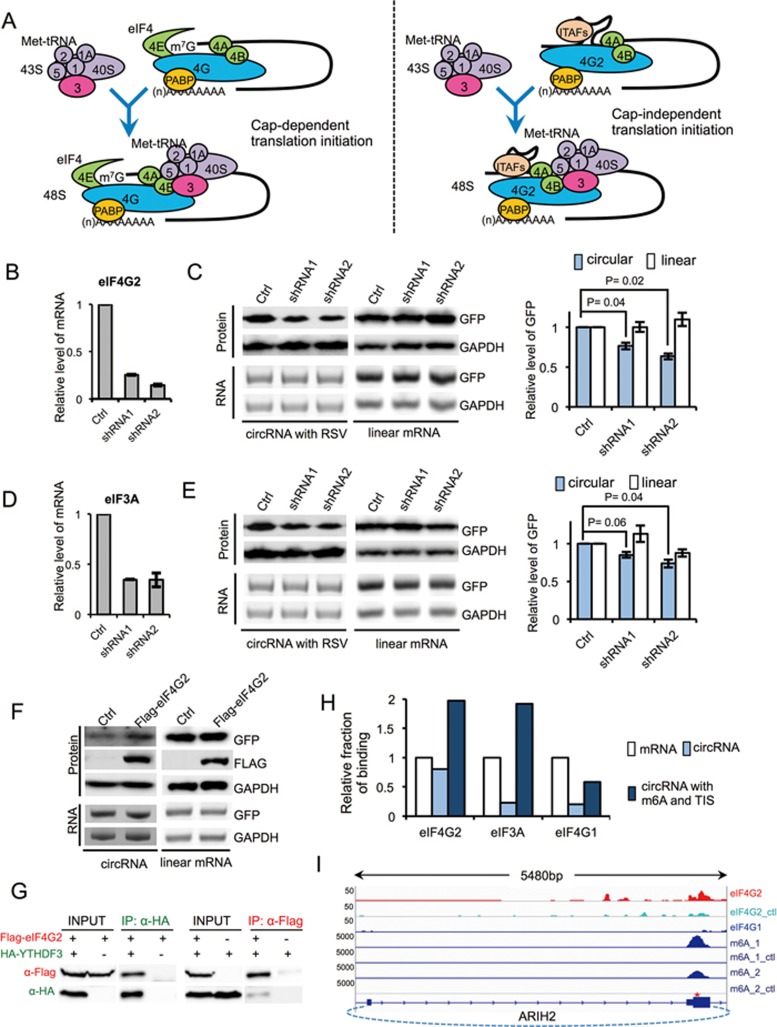
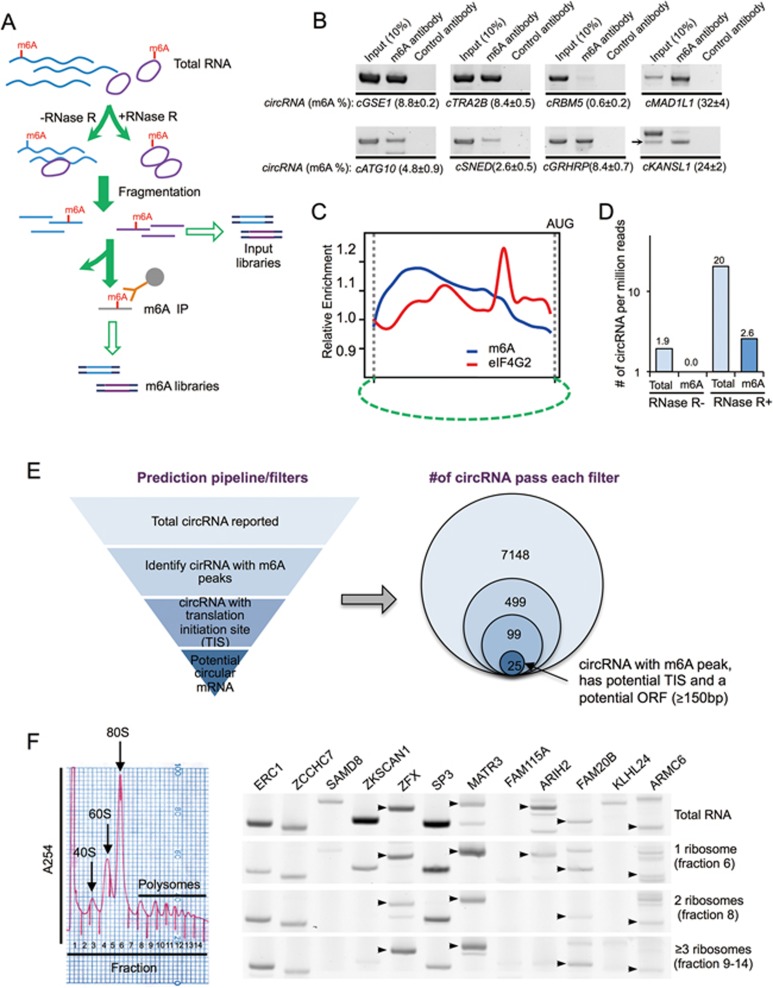
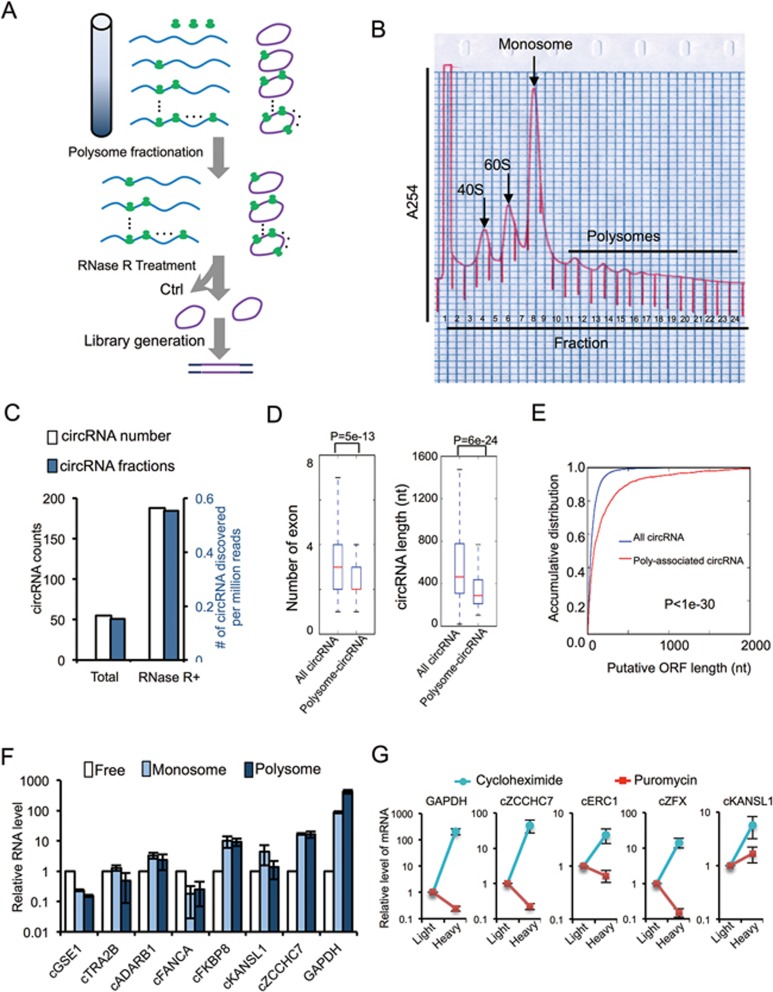
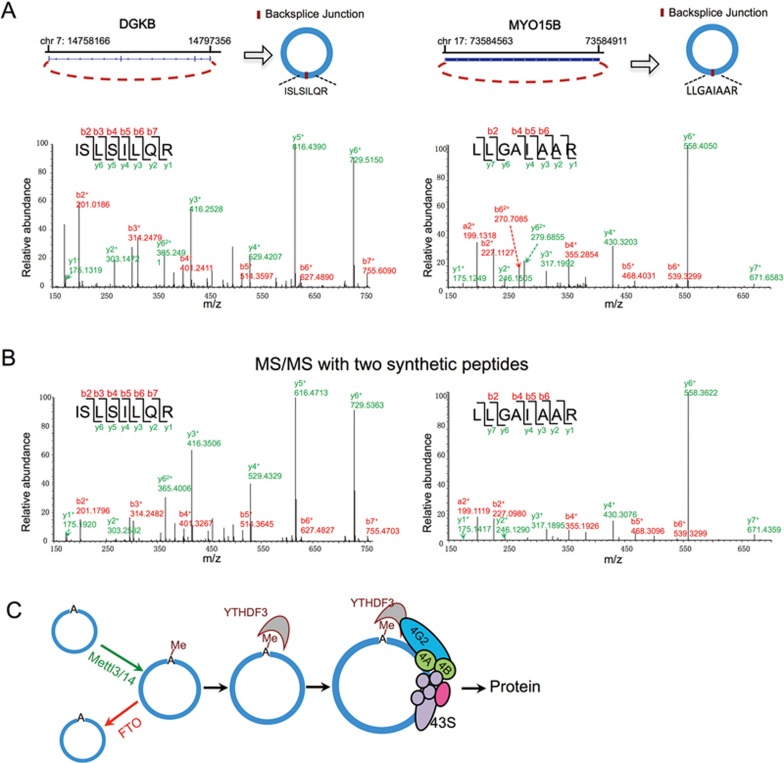
Extensive pre-mRNA back-splicing generates numerous circular RNAs (circRNAs) in human transcriptome. However, the biological functions of these circRNAs remain largely unclear. Here we report that N6-methyladenosine (m6A), the most abundant base modification of RNA, promotes efficient initiation of protein translation from circRNAs in human cells. We discover that consensus m6A motifs are enriched in circRNAs and a single m6A site is sufficient to drive translation initiation. This m6A-driven translation requires initiation factor eIF4G2 and m6A reader YTHDF3, and is enhanced by methyltransferase METTL3/14, inhibited by demethylase FTO, and upregulated upon heat shock. Further analyses through polysome profiling, computational prediction and mass spectrometry reveal that m6A-driven translation of circRNAs is widespread, with hundreds of endogenous circRNAs having translation potential. Our study expands the coding landscape of human transcriptome, and suggests a role of circRNA-derived proteins in cellular responses to environmental stress.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
