

HNF-4 participates in the hibernation-associated transcriptional regulation of the chipmunk hibernation-related protein gene
- PMID: 28281641
- PMCID: PMC5345028
- DOI: 10.1038/srep44279
HNF-4 participates in the hibernation-associated transcriptional regulation of the chipmunk hibernation-related protein gene
Abstract
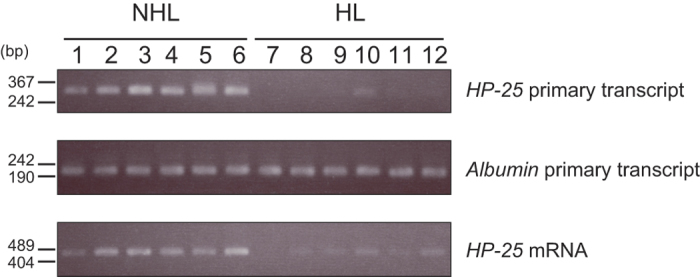
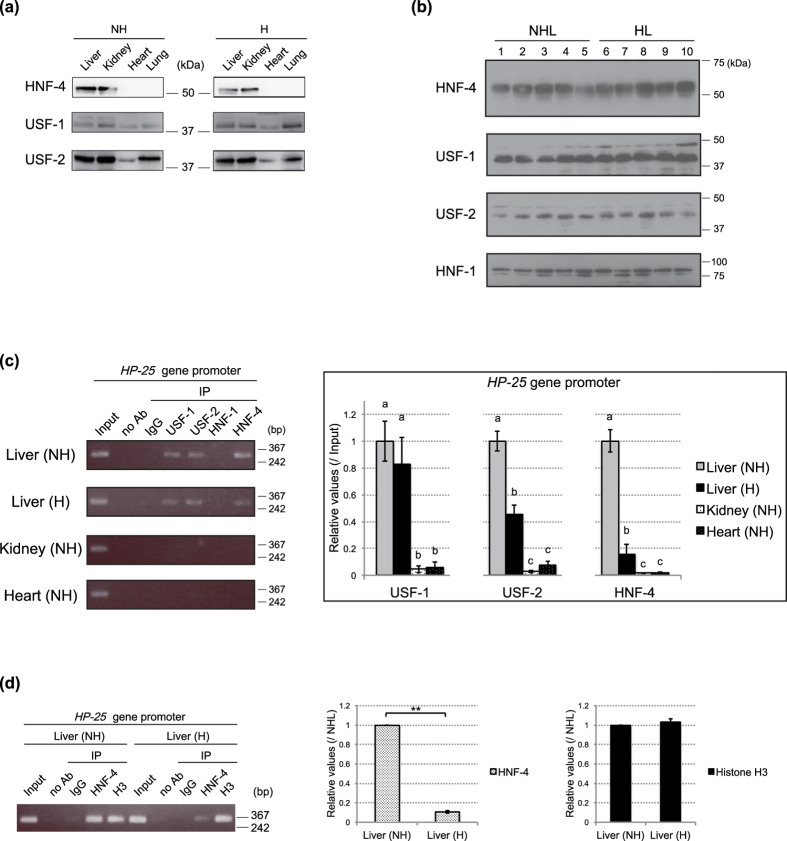
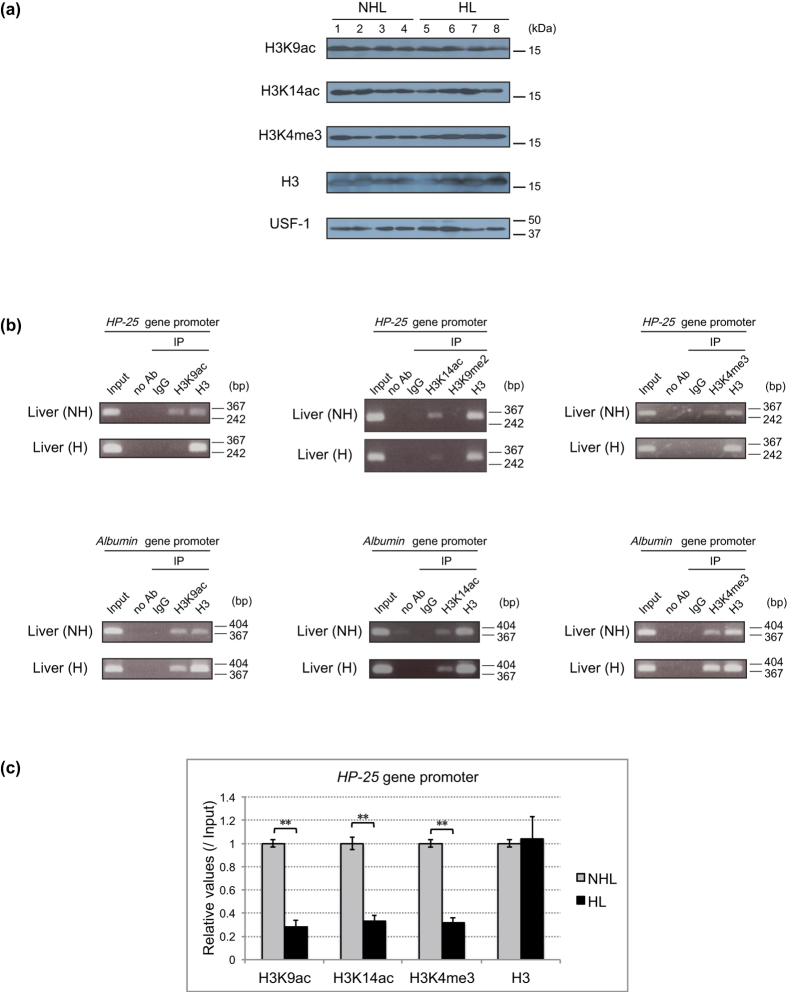
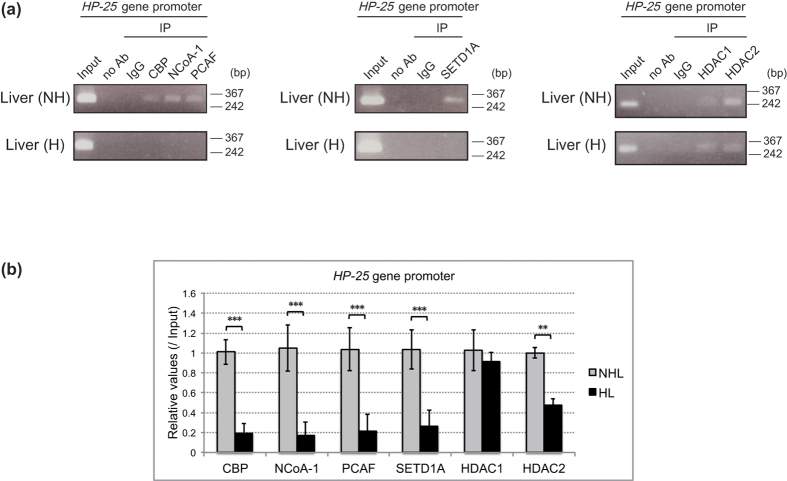
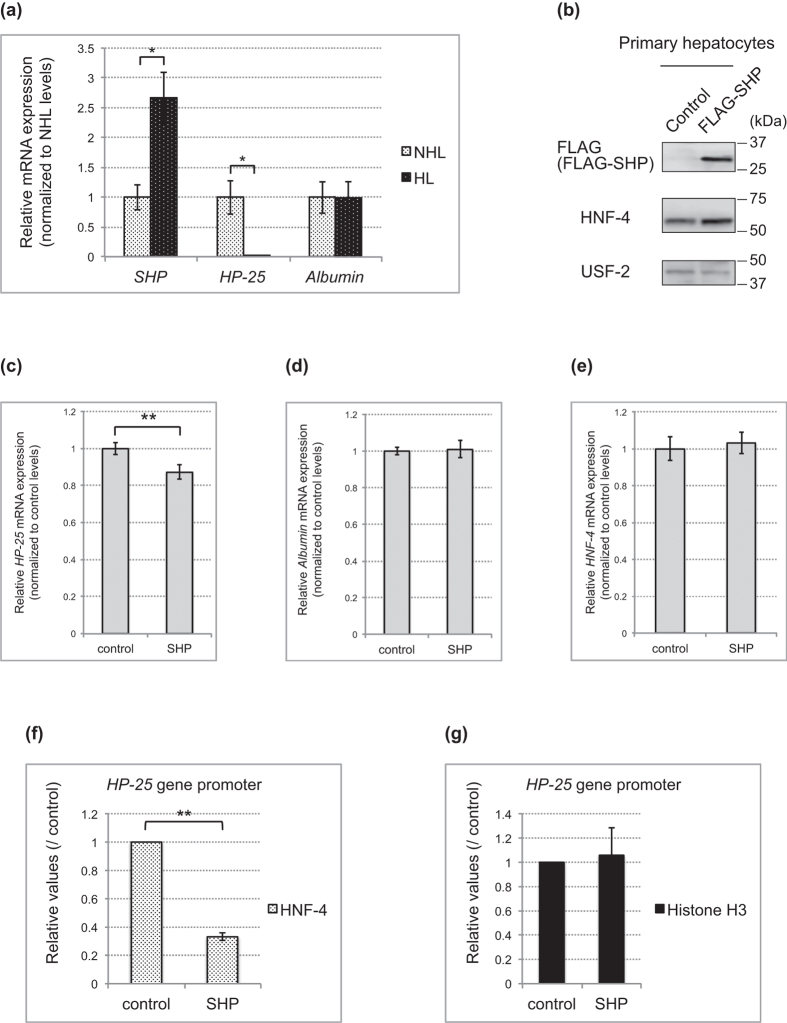
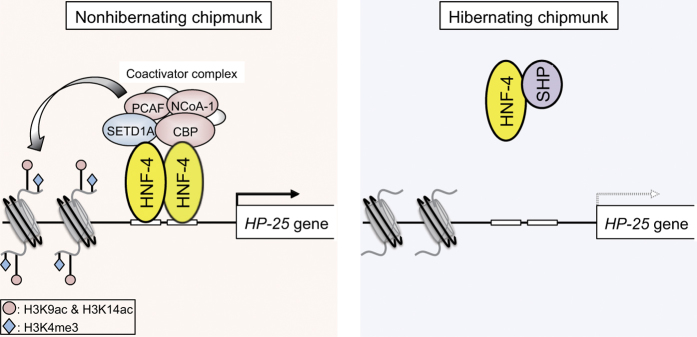
The chipmunk hibernation-related protein 25 (HP-25) is involved in the circannual control of hibernation in the brain. The liver-specific expression of the HP-25 gene is repressed in hibernating chipmunks under the control of endogenous circannual rhythms. However, the molecular mechanisms that differentially regulate the HP-25 gene during the nonhibernation and hibernation seasons are unknown. Here, we show that the hibernation-associated HP-25 expression is regulated epigenetically. Chromatin immunoprecipitation analyses revealed that significantly less hepatocyte nuclear receptor HNF-4 bound to the HP-25 gene promoter in the liver of hibernating chipmunks compared to nonhibernating chipmunks. Concurrently in the hibernating chipmunks, coactivators were dissociated from the promoter, and active transcription histone marks on the HP-25 gene promoter were lost. On the other hand, small heterodimer partner (SHP) expression was upregulated in the liver of hibernating chipmunks. Overexpressing SHP in primary hepatocytes prepared from nonhibernating chipmunks caused HNF-4 to dissociate from the HP-25 gene promoter, and reduced the HP-25 mRNA level. These results suggest that hibernation-related HP-25 expression is epigenetically regulated by the binding of HNF-4 to the HP-25 promoter, and that this binding might be modulated by SHP in hibernating chipmunks.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Wang L. C. H. & Lee T. F. Topor and Hibernation in Mammals: Metabolic, Physiological, and Biochemical Adaptations. In Comprehensive Physiology 507–532 (American Physiological Society, 2011).
-
- Kondo N. & Kondo J. Identification of novel blood proteins specific for mammalian hibernation. J. Biol. Chem. 267, 473–478 (1992). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
