

The transjugation machinery of Thermus thermophilus: Identification of TdtA, an ATPase involved in DNA donation
- PMID: 28282376
- PMCID: PMC5365140
- DOI: 10.1371/journal.pgen.1006669
The transjugation machinery of Thermus thermophilus: Identification of TdtA, an ATPase involved in DNA donation
Abstract
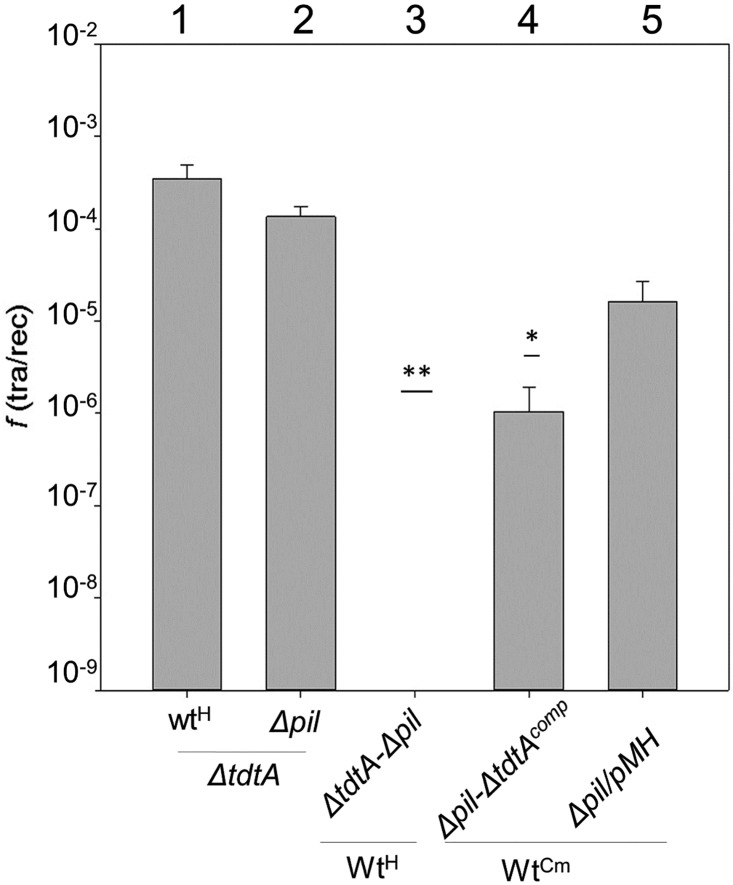
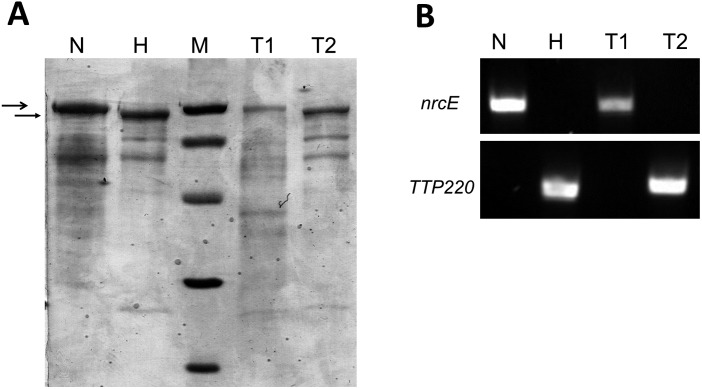
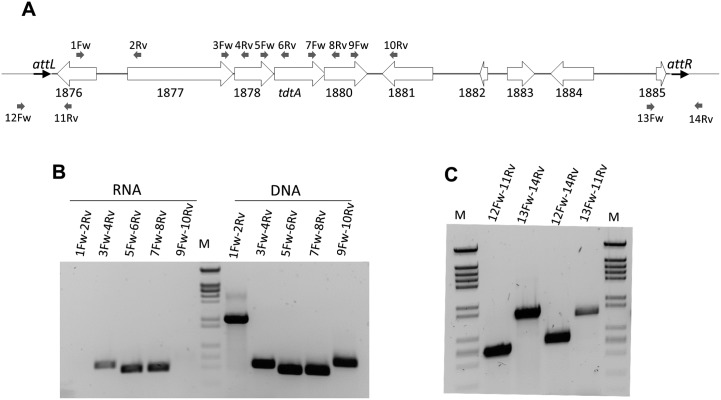
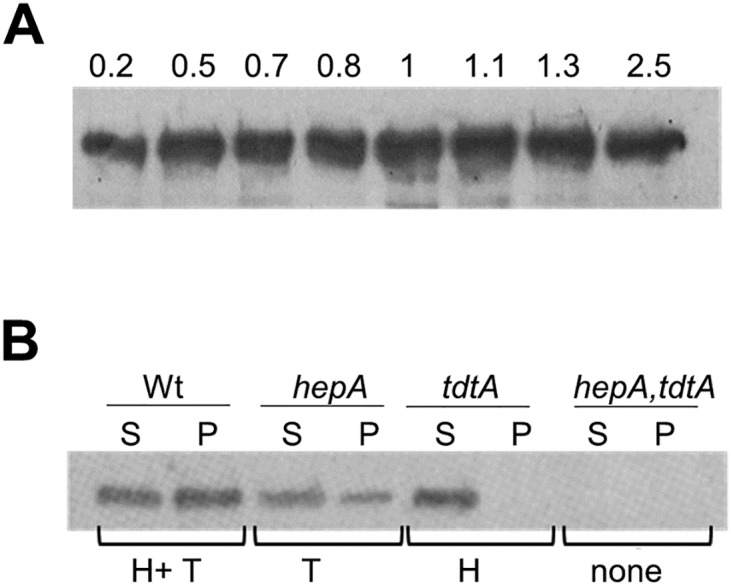
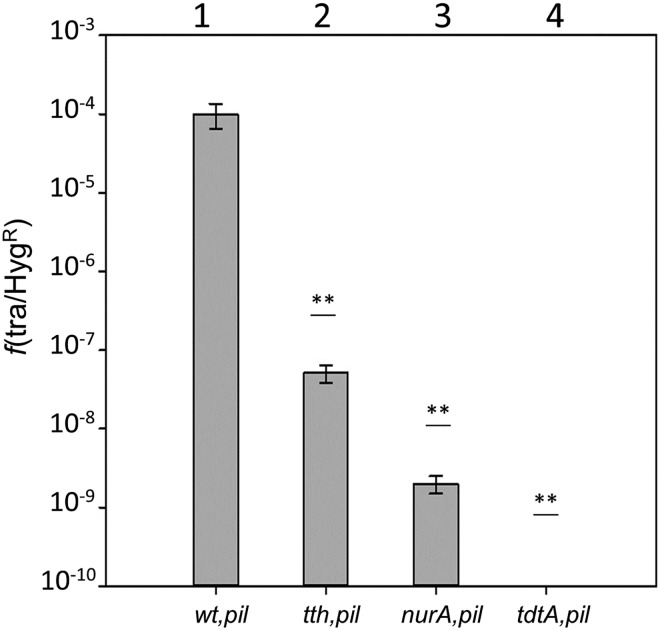
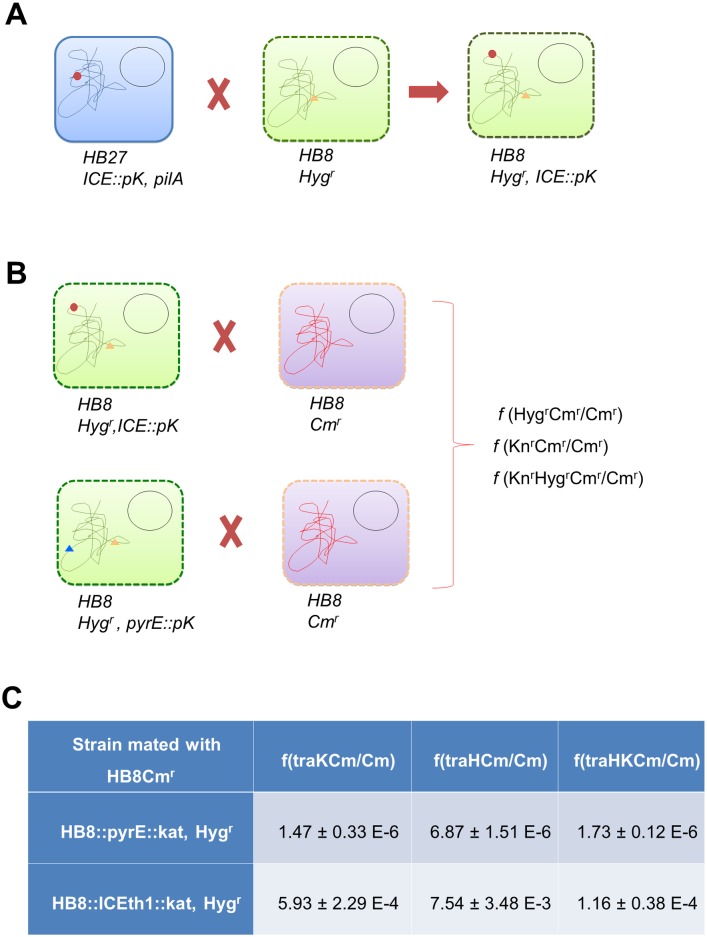
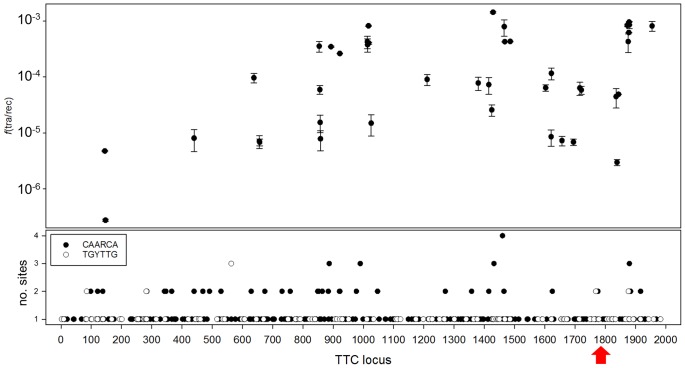
In addition to natural competence, some Thermus thermophilus strains show a high rate of DNA transfer via direct cell-to-cell contact. The process is bidirectional and follows a two-step model where the donor cell actively pushes out DNA and the recipient cell employs the natural competence system to take up the DNA, in a hybrid transformation-dependent conjugation process (transjugation). While the DNA uptake machinery is well known as in other bacterial species that undergo transformation, the pushing step of transjugation remains to be characterized. Here we have searched for hypothetical DNA translocases putatively involved in the pushing step of transjugation. Among candidates encoded by T. thermophilus HB27, the TdtA protein was found to be required for DNA pushing but not for DNA pulling during transjugation, without affecting other cellular processes. Purified TdtA shows ATPase activity and oligomerizes as hexamers with a central opening that can accommodate double-stranded DNA. The tdtA gene was found to belong to a mobile 14 kbp-long DNA element inserted within the 3' end of a tRNA gene, flanked by 47 bp direct repeats. The insertion also encoded a homolog of bacteriophage site-specific recombinases and actively self-excised from the chromosome at high frequency to form an apparently non-replicative circular form. The insertion also encoded a type II restriction endonuclease and a NurA-like nuclease, whose activities were required for efficient transjugation. All these data support that TdtA belongs to a new type of Integrative and Conjugative Element which promotes the generalized and efficient transfer of genetic traits that could facilitate its co-selection among bacterial populations.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures

References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
