

Trypanosoma brucei metabolism is under circadian control
- PMID: 28288095
- PMCID: PMC5398093
- DOI: 10.1038/nmicrobiol.2017.32
Trypanosoma brucei metabolism is under circadian control
Abstract
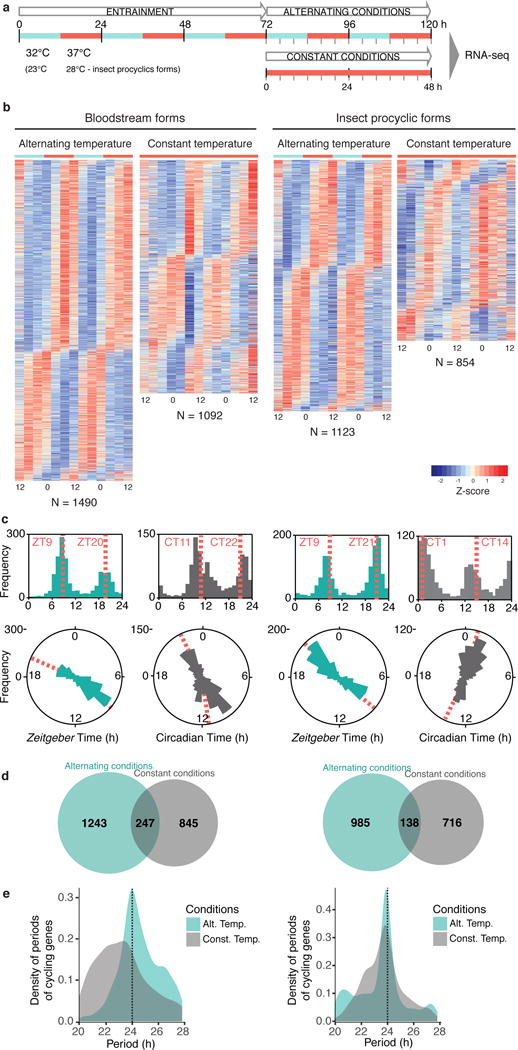
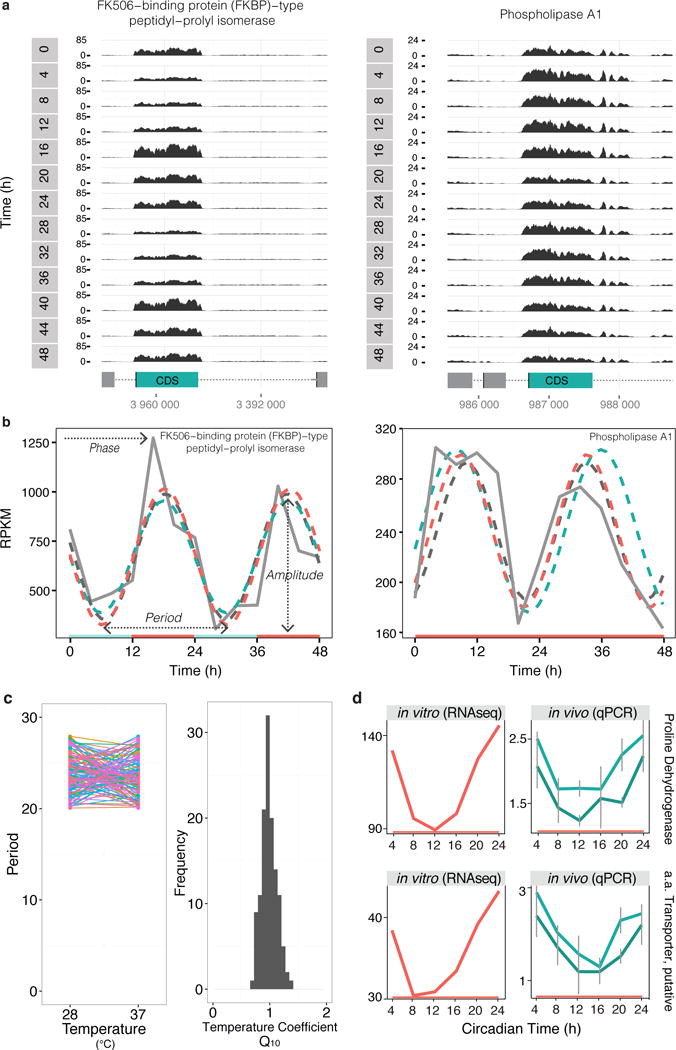
The Earth's rotation forced life to evolve under cyclic day and night environmental changes. To anticipate such daily cycles, prokaryote and eukaryote free-living organisms evolved intrinsic clocks that regulate physiological and behavioural processes. Daily rhythms have been observed in organisms living within hosts, such as parasites. Whether parasites have intrinsic molecular clocks or whether they simply respond to host rhythmic physiological cues remains unknown. Here, we show that Trypanosoma brucei, the causative agent of human sleeping sickness, has an intrinsic circadian clock that regulates its metabolism in two different stages of the life cycle. We found that, in vitro, ∼10% of genes in T. brucei are expressed with a circadian rhythm. The maximum expression of these genes occurs at two different phases of the day and may depend on a post-transcriptional mechanism. Circadian genes are enriched in cellular metabolic pathways and coincide with two peaks of intracellular adenosine triphosphate concentration. Moreover, daily changes in the parasite population lead to differences in suramin sensitivity, a drug commonly used to treat this infection. These results demonstrate that parasites have an intrinsic circadian clock that is independent of the host, and which regulates parasite biology throughout the day.
Conflict of interest statement
The authors declare no competing financial interests.
Figures



References
-
- Balsalobre A, Damiola F, Schibler U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell. 1998;93:929–937. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
