

Disruption of the ATXN1-CIC complex causes a spectrum of neurobehavioral phenotypes in mice and humans
- PMID: 28288114
- PMCID: PMC5374026
- DOI: 10.1038/ng.3808
Disruption of the ATXN1-CIC complex causes a spectrum of neurobehavioral phenotypes in mice and humans
Abstract
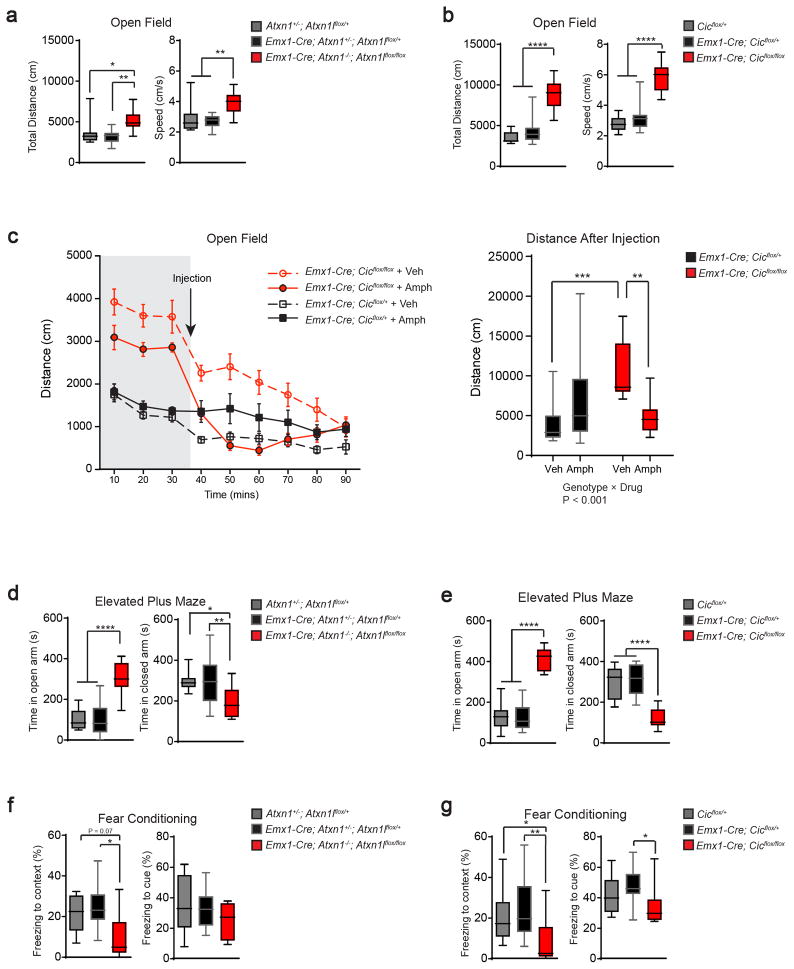
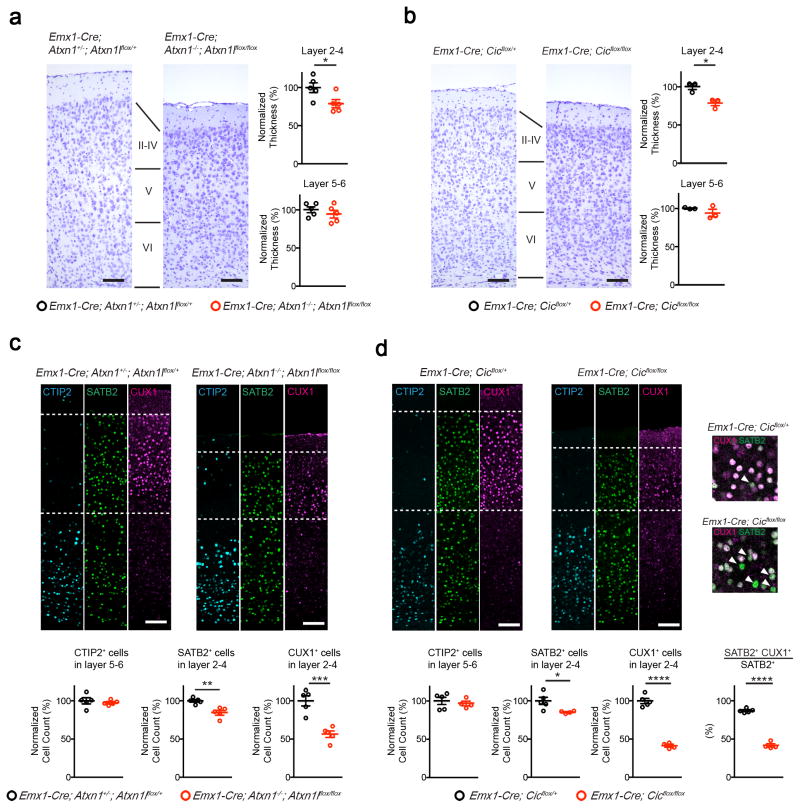
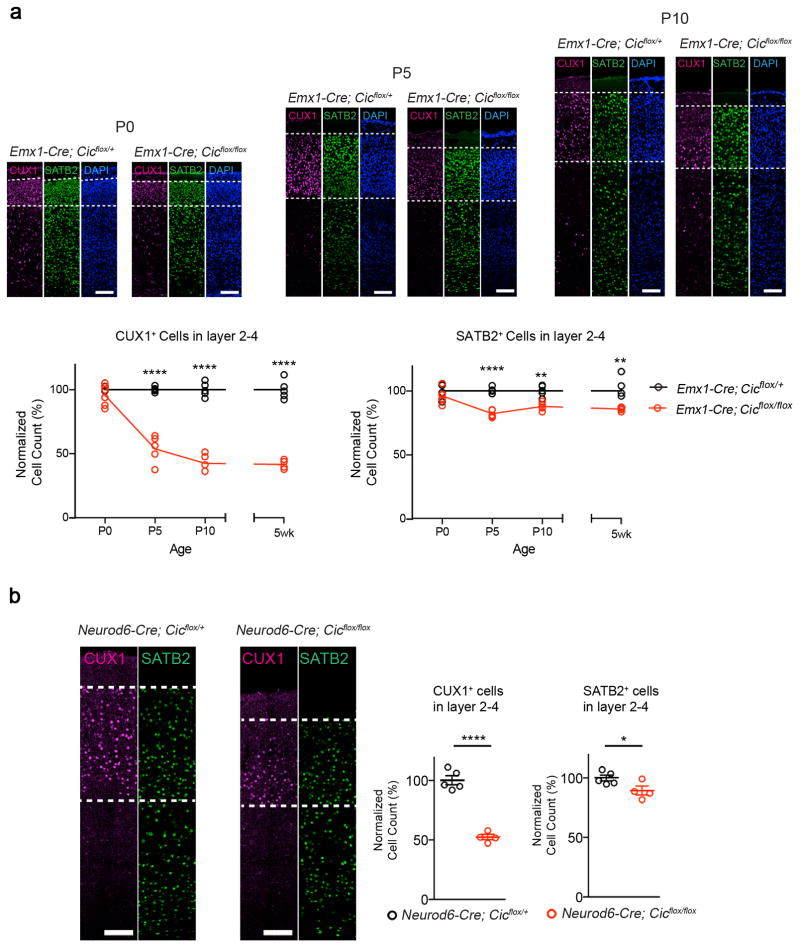
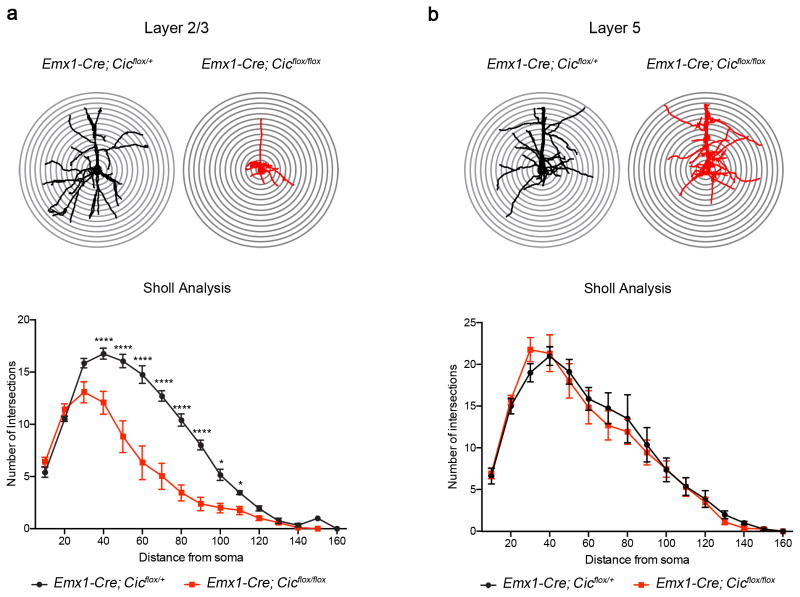
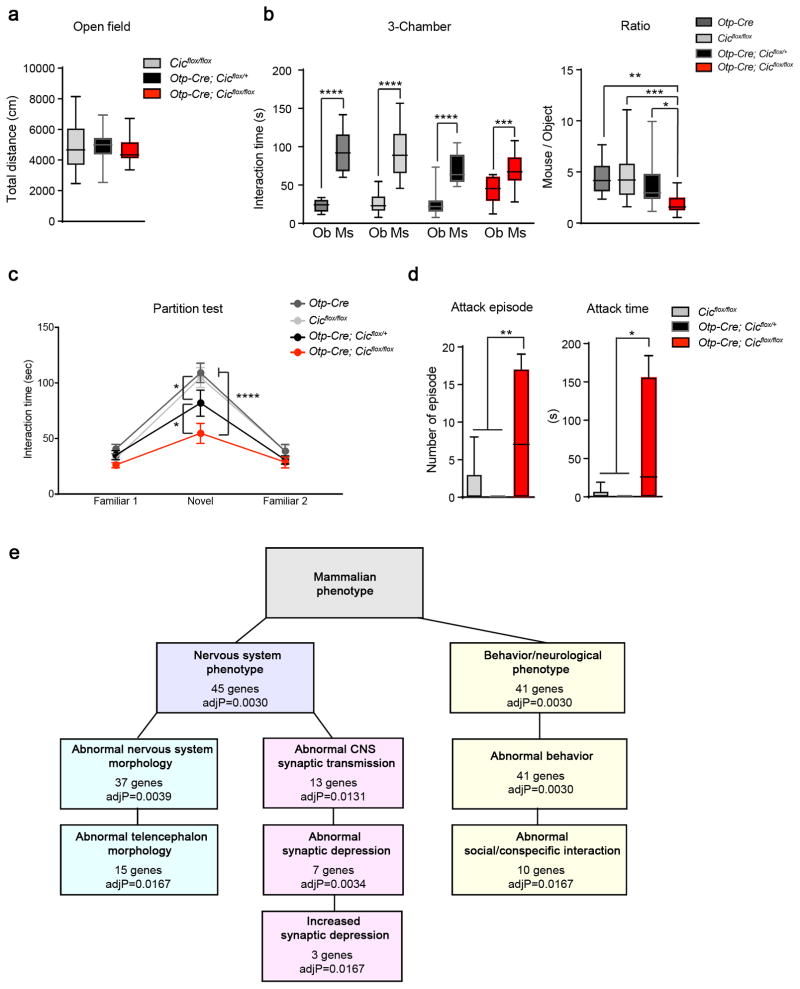
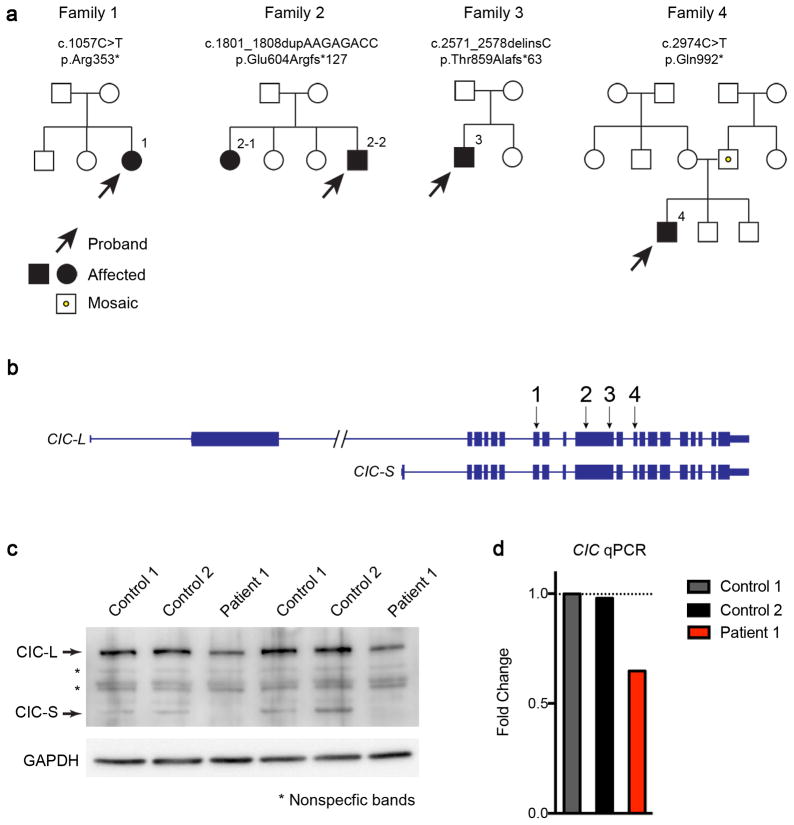
Gain-of-function mutations in some genes underlie neurodegenerative conditions, whereas loss-of-function mutations in the same genes have distinct phenotypes. This appears to be the case with the protein ataxin 1 (ATXN1), which forms a transcriptional repressor complex with capicua (CIC). Gain of function of the complex leads to neurodegeneration, but ATXN1-CIC is also essential for survival. We set out to understand the functions of the ATXN1-CIC complex in the developing forebrain and found that losing this complex results in hyperactivity, impaired learning and memory, and abnormal maturation and maintenance of upper-layer cortical neurons. We also found that CIC activity in the hypothalamus and medial amygdala modulates social interactions. Informed by these neurobehavioral features in mouse mutants, we identified five individuals with de novo heterozygous truncating mutations in CIC who share similar clinical features, including intellectual disability, attention deficit/hyperactivity disorder (ADHD), and autism spectrum disorder. Our study demonstrates that loss of ATXN1-CIC complexes causes a spectrum of neurobehavioral phenotypes.
Conflict of interest statement
None declared.
Figures
References
-
- Orr HT, et al. Expansion of an unstable trinucleotide CAG repeat in spinocerebellar ataxia type 1. Nat Genet. 1993;4:221–6. - PubMed
-
- Burright EN, et al. SCA1 transgenic mice: a model for neurodegeneration caused by an expanded CAG trinucleotide repeat. Cell. 1995;82:937–48. - PubMed
-
- Watase K, et al. A long CAG repeat in the mouse Sca1 locus replicates SCA1 features and reveals the impact of protein solubility on selective neurodegeneration. Neuron. 2002;34:905–19. - PubMed
-
- Asher M, Johnson A, Zecevic B, Pease D, Cvetanovic M. Ataxin-1 regulates proliferation of hippocampal neural precursors. Neuroscience. 2016 - PubMed
MeSH terms
Substances
Grants and funding
- R01 HD083809/HD/NICHD NIH HHS/United States
- S10 OD016167/OD/NIH HHS/United States
- F32 NS083091/NS/NINDS NIH HHS/United States
- U54 HD083092/HD/NICHD NIH HHS/United States
- R37 NS022920/NS/NINDS NIH HHS/United States
- HHMI/Howard Hughes Medical Institute/United States
- P01 HD067244/HD/NICHD NIH HHS/United States
- R37 NS027699/NS/NINDS NIH HHS/United States
- R01 NS027699/NS/NINDS NIH HHS/United States
- UM1 HG008898/HG/NHGRI NIH HHS/United States
- R01 NS089664/NS/NINDS NIH HHS/United States
- R01 HD081216/HD/NICHD NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
