

The Pseudomonas aeruginosa PrrF Small RNAs Regulate Iron Homeostasis during Acute Murine Lung Infection
- PMID: 28289146
- PMCID: PMC5400841
- DOI: 10.1128/IAI.00764-16
The Pseudomonas aeruginosa PrrF Small RNAs Regulate Iron Homeostasis during Acute Murine Lung Infection
Abstract
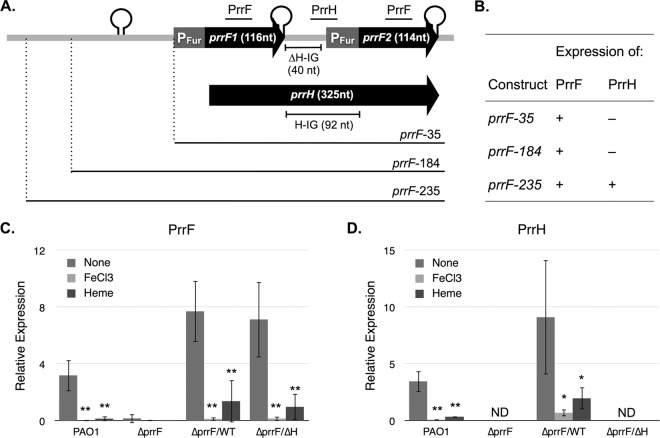
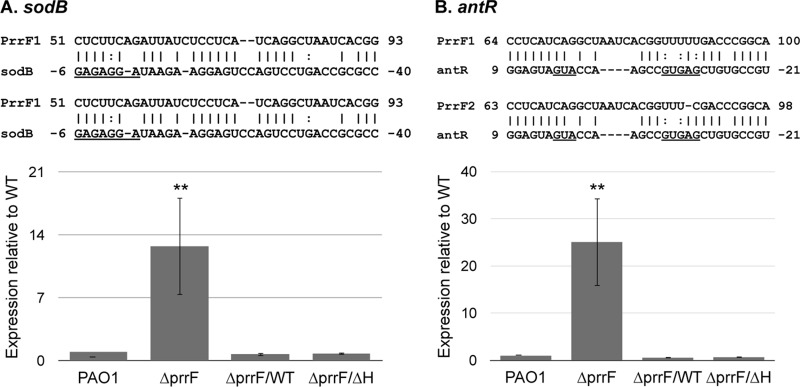
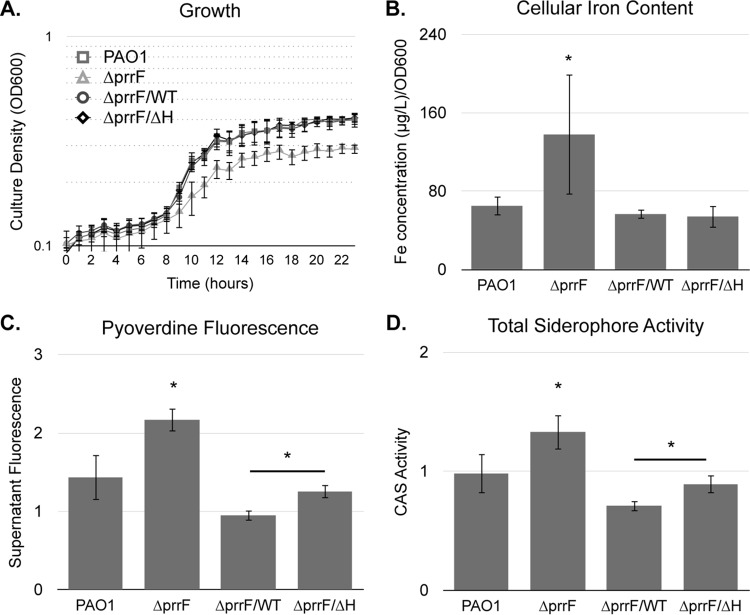
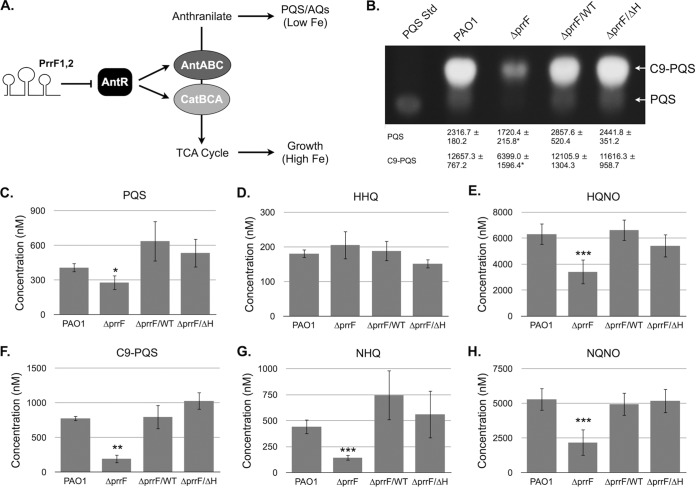
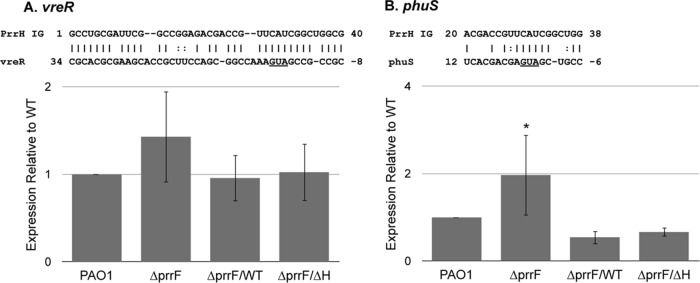
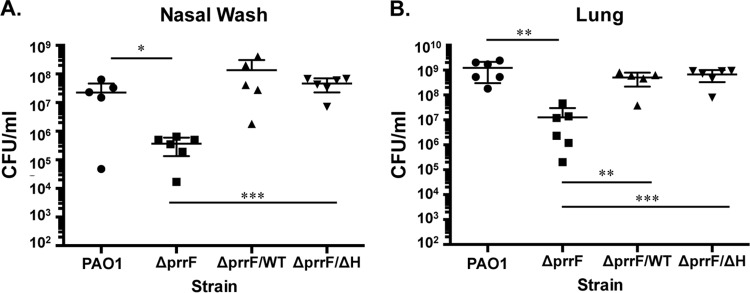
Pseudomonas aeruginosa is a Gram-negative opportunistic pathogen that requires iron for virulence. Iron homeostasis is maintained in part by the PrrF1 and PrrF2 small RNAs (sRNAs), which block the expression of iron-containing proteins under iron-depleted conditions. The PrrF sRNAs also promote the production of the Pseudomonas quinolone signal (PQS), a quorum sensing molecule that activates the expression of several virulence genes. The tandem arrangement of the prrF genes allows for expression of a third sRNA, PrrH, which is predicted to regulate gene expression through its unique sequence derived from the prrF1-prrF2 intergenic (IG) sequence (the PrrHIG sequence). Previous studies showed that the prrF locus is required for acute lung infection. However, the individual functions of the PrrF and PrrH sRNAs were not determined. Here, we describe a system for differentiating PrrF and PrrH functions by deleting the PrrHIG sequence [prrF(ΔHIG)]. Our analyses of this construct indicate that the PrrF sRNAs, but not PrrH, are required for acute lung infection by P. aeruginosa Moreover, we show that the virulence defect of the ΔprrF1-prrF2 mutant is due to decreased bacterial burden during acute lung infection. In vivo analysis of gene expression in lung homogenates shows that PrrF-mediated regulation of genes for iron-containing proteins is disrupted in the ΔprrF1-prrF2 mutant during infection, while the expression of genes that mediate PrrF-regulated PQS production are not affected by prrF deletion in vivo Combined, these studies demonstrate that regulation of iron utilization plays a critical role in P. aeruginosa's ability to survive during infection.
Keywords: PQS; PrrF; PrrH; Pseudomonas aeruginosa; iron regulation; sRNA; small RNA.
Copyright © 2017 American Society for Microbiology.
Figures
Similar articles
-
The prrF-encoded small regulatory RNAs are required for iron homeostasis and virulence of Pseudomonas aeruginosa.Infect Immun. 2015 Mar;83(3):863-75. doi: 10.1128/IAI.02707-14. Epub 2014 Dec 15. Infect Immun. 2015. PMID: 25510881 Free PMC article.
-
The Pseudomonas aeruginosa PrrF1 and PrrF2 Small Regulatory RNAs Promote 2-Alkyl-4-Quinolone Production through Redundant Regulation of the antR mRNA.J Bacteriol. 2018 Apr 24;200(10):e00704-17. doi: 10.1128/JB.00704-17. Print 2018 May 15. J Bacteriol. 2018. PMID: 29507088 Free PMC article.
-
The heme-responsive PrrH sRNA regulates Pseudomonas aeruginosa pyochelin gene expression.mSphere. 2023 Oct 24;8(5):e0039223. doi: 10.1128/msphere.00392-23. Epub 2023 Oct 6. mSphere. 2023. PMID: 37800921 Free PMC article.
-
How we learnt about iron acquisition in Pseudomonas aeruginosa: a series of very fortunate events.Biometals. 2007 Jun;20(3-4):587-601. doi: 10.1007/s10534-006-9067-2. Epub 2006 Dec 22. Biometals. 2007. PMID: 17186376 Review.
-
Relationship between Pyochelin and Pseudomonas Quinolone Signal in Pseudomonas aeruginosa: A Direction for Future Research.Int J Mol Sci. 2024 Aug 7;25(16):8611. doi: 10.3390/ijms25168611. Int J Mol Sci. 2024. PMID: 39201297 Free PMC article. Review.
Cited by
-
ECF Sigma Factor HxuI Is Critical for In Vivo Fitness of Pseudomonas aeruginosa during Infection.Microbiol Spectr. 2022 Feb 23;10(1):e0162021. doi: 10.1128/spectrum.01620-21. Epub 2022 Jan 19. Microbiol Spectr. 2022. PMID: 35044199 Free PMC article.
-
Proteomic Analysis of the Pseudomonas aeruginosa Iron Starvation Response Reveals PrrF Small Regulatory RNA-Dependent Iron Regulation of Twitching Motility, Amino Acid Metabolism, and Zinc Homeostasis Proteins.J Bacteriol. 2019 May 22;201(12):e00754-18. doi: 10.1128/JB.00754-18. Print 2019 Jun 15. J Bacteriol. 2019. PMID: 30962354 Free PMC article.
-
The Pseudomonas aeruginosa PhuS proximal ligand His-209 triggers a conformational switch in function from DNA binding to heme transfer.J Biol Chem. 2025 May;301(5):108440. doi: 10.1016/j.jbc.2025.108440. Epub 2025 Mar 22. J Biol Chem. 2025. PMID: 40122173 Free PMC article.
-
Static Growth Promotes PrrF and 2-Alkyl-4(1H)-Quinolone Regulation of Type VI Secretion Protein Expression in Pseudomonas aeruginosa.J Bacteriol. 2020 Nov 19;202(24):e00416-20. doi: 10.1128/JB.00416-20. Print 2020 Nov 19. J Bacteriol. 2020. PMID: 33020221 Free PMC article.
-
Small RNA Regulation of Virulence in Pathogenic Escherichia coli.Front Cell Infect Microbiol. 2021 Jan 27;10:622202. doi: 10.3389/fcimb.2020.622202. eCollection 2020. Front Cell Infect Microbiol. 2021. PMID: 33585289 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
