

The schizophrenia- and autism-associated gene, transcription factor 4 regulates the columnar distribution of layer 2/3 prefrontal pyramidal neurons in an activity-dependent manner
- PMID: 28289282
- PMCID: PMC5599320
- DOI: 10.1038/mp.2017.37
The schizophrenia- and autism-associated gene, transcription factor 4 regulates the columnar distribution of layer 2/3 prefrontal pyramidal neurons in an activity-dependent manner
Abstract
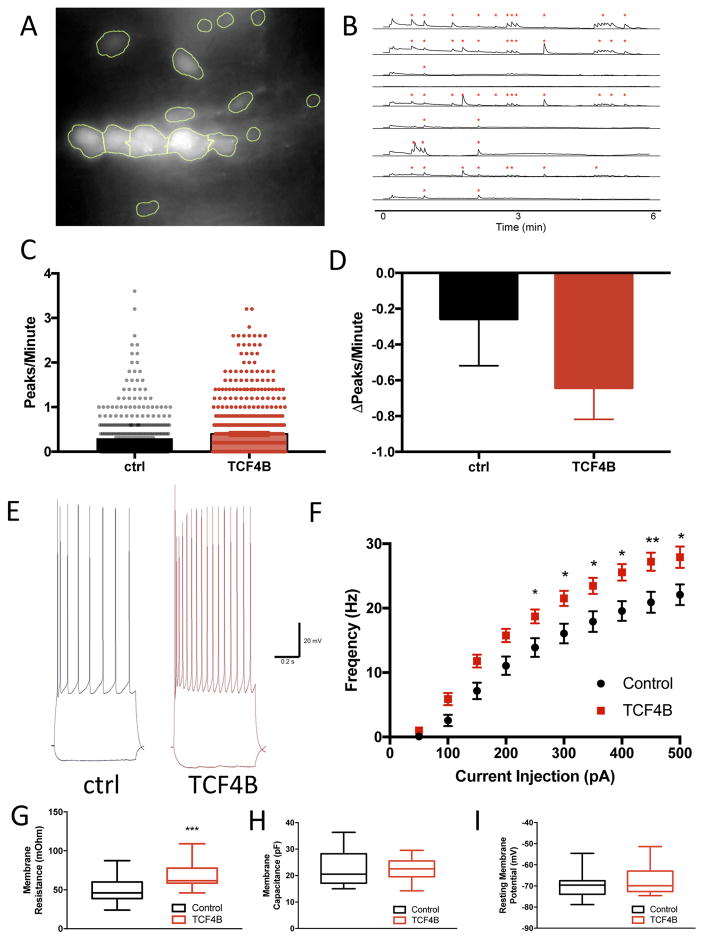
Disruption of the laminar and columnar organization of the brain is implicated in several psychiatric disorders. Here, we show in utero gain-of-function of the psychiatric risk gene transcription factor 4 (TCF4) severely disrupts the columnar organization of medial prefrontal cortex (mPFC) in a transcription- and activity-dependent manner. This morphological phenotype was rescued by co-expression of TCF4 plus calmodulin in a calcium-dependent manner and by dampening neuronal excitability through co-expression of an inwardly rectifying potassium channel (Kir2.1). For we believe the first time, we show that N-methyl-d-aspartate (NMDA) receptor-dependent Ca2+ transients are instructive to minicolumn organization because Crispr/Cas9-mediated mutation of NMDA receptors rescued TCF4-dependent morphological phenotypes. Furthermore, we demonstrate that the transcriptional regulation by the psychiatric risk gene TCF4 enhances NMDA receptor-dependent early network oscillations. Our novel findings indicate that TCF4-dependent transcription directs the proper formation of prefrontal cortical minicolumns by regulating the expression of genes involved in early spontaneous neuronal activity, and thus our results provides insights into potential pathophysiological mechanisms of TCF4-associated psychiatric disorders.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Casanova MF, Buxhoeveden DP, Switala AE, Roy E. Minicolumnar pathology in autism. Neurology. 2002;58:428–432. - PubMed
-
- Casanova MF, Buxhoeveden DP, Cohen M, Switala AE, Roy EL. Minicolumnar pathology in dyslexia. Ann Neurol. 2002;52:108–110. - PubMed
-
- Casanova MF, de Zeeuw L, Switala A, Kreczmanski P, Korr H, Ulfig N, et al. Mean cell spacing abnormalities in the neocortex of patients with schizophrenia. Psychiatry Research. 2005;133:1–12. - PubMed
-
- Gleeson JG. Classical lissencephaly and double cortex (subcortical band heterotopia): LIS1 and doublecortin. Curr Opin Neurol. 2000;13:121–125. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
