

PTEN proteoforms in biology and disease
- PMID: 28289760
- PMCID: PMC11107534
- DOI: 10.1007/s00018-017-2500-6
PTEN proteoforms in biology and disease
Abstract
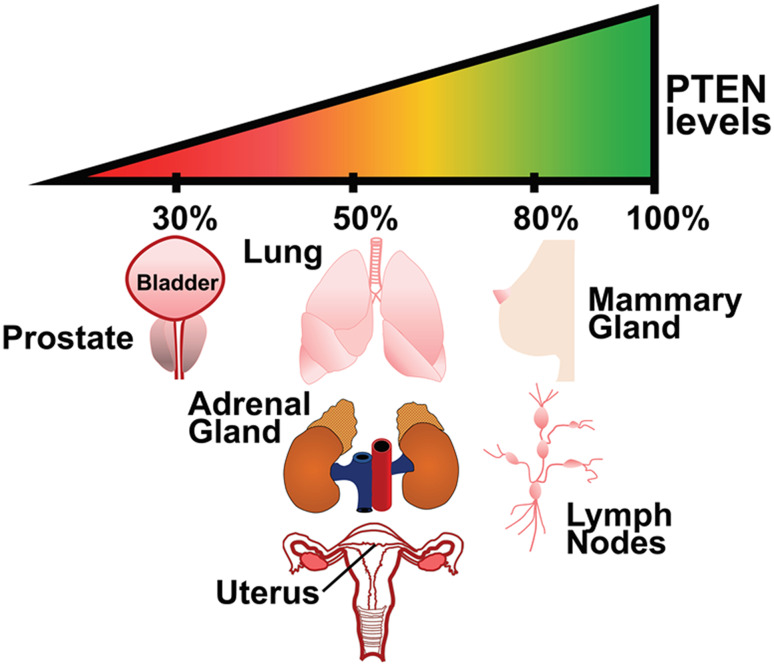
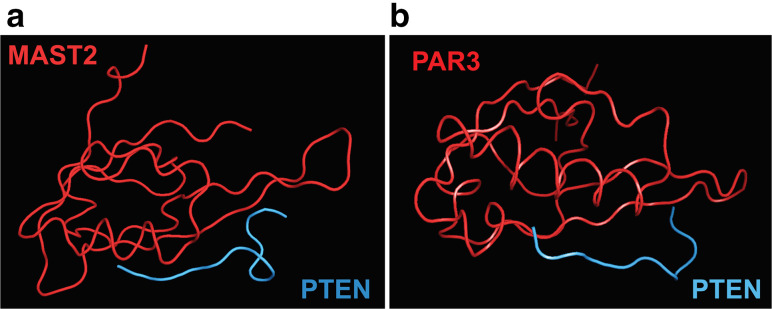
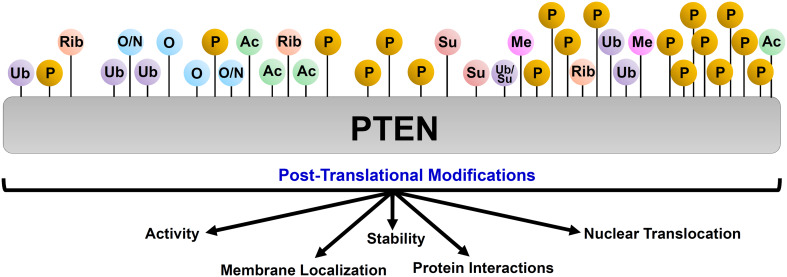
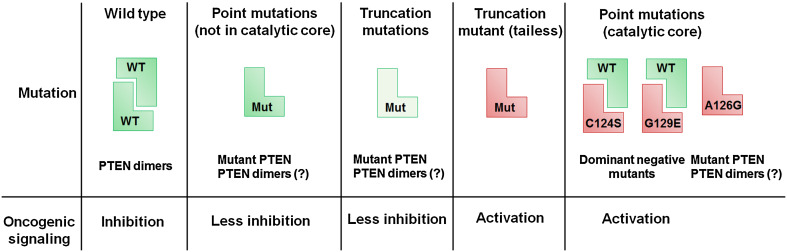
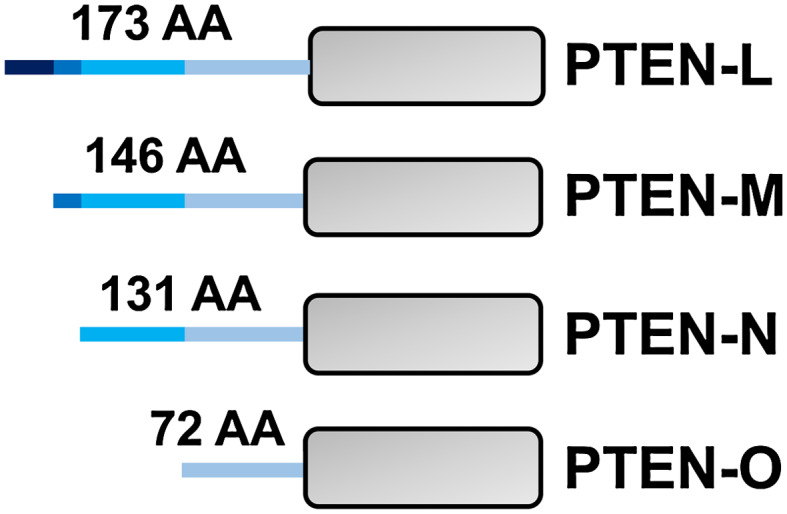
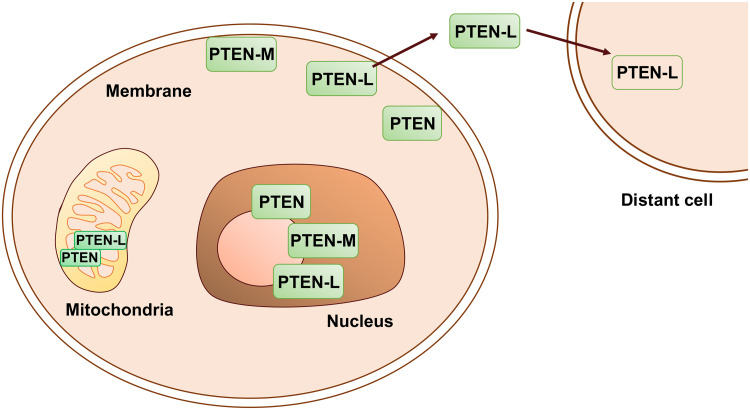
Proteoforms are specific molecular forms of protein products arising from a single gene that possess different structures and different functions. Therefore, a single gene can produce a large repertoire of proteoforms by means of allelic variations (mutations, indels, SNPs), alternative splicing and other pre-translational mechanisms, post-translational modifications (PTMs), conformational dynamics, and functioning. Resulting proteoforms that have different sizes, alternative splicing patterns, sets of post-translational modifications, protein-protein interactions, and protein-ligand interactions, might dramatically increase the functionality of the encoded protein. Herein, we have interrogated the tumor suppressor PTEN for its proteoforms and find that this protein exists in multiple forms with distinct functions and sub-cellular localizations. Furthermore, the levels of each PTEN proteoform in a given cell may affect its biological function. Indeed, the paradigm of the continuum model of tumor suppression by PTEN can be better explained by the presence of a continuum of PTEN proteoforms, diversity, and levels of which are associated with pathological outcomes than simply by the different roles of mutations in the PTEN gene. Consequently, understanding the mechanisms underlying the dysregulation of PTEN proteoforms by several genomic and non-genomic mechanisms in cancer and other diseases is imperative. We have identified different PTEN proteoforms, which control various aspects of cellular function and grouped them into three categories of intrinsic, function-induced, and inducible proteoforms. A special emphasis is given to the inducible PTEN proteoforms that are produced due to alternative translational initiation. The novel finding that PTEN forms dimers with biological implications supports the notion that PTEN proteoform-proteoform interactions may play hitherto unknown roles in cellular homeostasis and in pathogenic settings, including cancer. These PTEN proteoforms with unique properties and functionalities offer potential novel therapeutic opportunities in the treatment of various cancers and other diseases.
Keywords: Alternative translational initiation; PTEN; Post-translational modifications; Proteoforms.
Figures

References
-
- Li J, Yen C, Liaw D, Podsypanina K, Bose S, Wang SI, Puc J, Miliaresis C, Rodgers L, McCombie R, Bigner SH, Giovanella BC, Ittmann M, Tycko B, Hibshoosh H, Wigler MH, Parsons R. PTEN, a putative protein tyrosine phosphatase gene mutated in human brain, breast, and prostate cancer. Science. 1997;275(5308):1943–1947. doi: 10.1126/science.275.5308.1943. - DOI - PubMed
-
- Steck PA, Pershouse MA, Jasser SA, Yung WKA, Lin H, Ligon AH, Langford LA, Baumgard ML, Hattier T, Davis T, Frye C, Hu R, Swedlund B, Teng DHF, Tavtigian SV. Identification of a candidate tumour suppressor gene, MMAC1, at chromosome 10q23.3 that is mutated in multiple advanced cancers. Nat Genet. 1997;15(4):356–362. doi: 10.1038/ng0497-356. - DOI - PubMed
-
- Ramaswamy S, Nakamura N, Vazquez F, Batt DB, Perera S, Roberts TM, Sellers WR. Regulation of G(1) progression by the PTEN tumor suppressor protein is linked to inhibition of the phosphatidylinositol 3-kinase/Akt pathway. Proc Natl Acad Sci U S A. 1999;96(5):2110–2115. doi: 10.1073/pnas.96.5.2110. - DOI - PMC - PubMed
-
- You DW, Xin JP, Volk A, Wei W, Schmidt R, Scurti G, Nand S, Breuer EK, Kuo PC, Breslin P, Kini AR, Nishimura MI, Zeleznik-Le NJ, Zhang JW. FAK mediates a compensatory survival signal parallel to PI3K-AKT in PTEN-Null T-ALL cells. Cell Rep. 2015;10(12):2055–2068. doi: 10.1016/j.celrep.2015.02.056. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
