

High binding affinity of repressor IolR avoids costs of untimely induction of myo-inositol utilization by Salmonella Typhimurium
- PMID: 28290506
- PMCID: PMC5349611
- DOI: 10.1038/srep44362
High binding affinity of repressor IolR avoids costs of untimely induction of myo-inositol utilization by Salmonella Typhimurium
Abstract
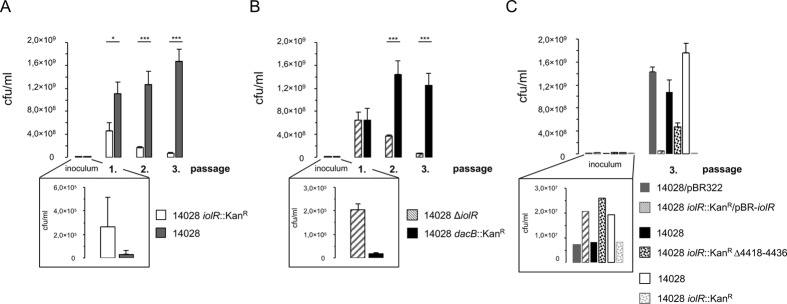
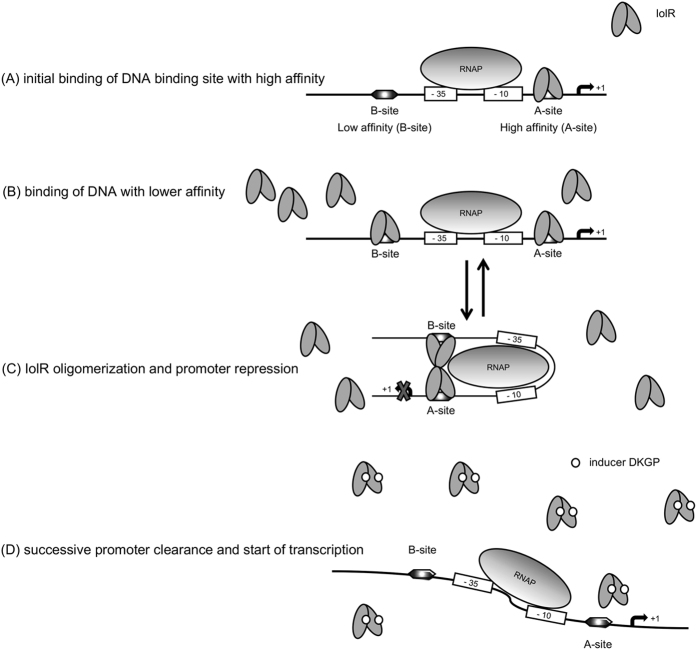
Growth of Salmonella enterica serovar Typhimurium strain 14028 with myo-inositol (MI) is characterized by a bistable phenotype that manifests with an extraordinarily long (34 h) and variable lag phase. When cells were pre-grown in minimal medium with MI, however, the lag phase shortened drastically to eight hours, and to six hours in the absence of the regulator IolR. To unravel the molecular mechanism behind this phenomenon, we investigated this repressor in more detail. Flow cytometry analysis of the iolR promoter at a single cell level demonstrated bistability of its transcriptional activation. Electrophoretic mobility shift assays were used to narrow the potential binding region of IolR and identified at least two binding sites in most iol gene promoters. Surface plasmon resonance spectroscopy quantified IolR binding and indicated its putative oligomerization and high binding affinity towards specific iol gene promoters. In competitive assays, the iolR deletion mutant, in which iol gene repression is abolished, showed a severe growth disadvantage of ~15% relative to the parental strain in rich medium. We hypothesize that the strong repression of iol gene transcription is required to maintain a balance between metabolic flexibility and fitness costs, which follow the inopportune induction of an unusual metabolic pathway.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Characterization of the myo-inositol utilization island of Salmonella enterica serovar Typhimurium.J Bacteriol. 2009 Jan;191(2):545-54. doi: 10.1128/JB.01253-08. Epub 2008 Nov 14. J Bacteriol. 2009. PMID: 19011032 Free PMC article.
-
Bistability in myo-inositol utilization by Salmonella enterica serovar Typhimurium.J Bacteriol. 2011 Mar;193(6):1427-35. doi: 10.1128/JB.00043-10. Epub 2011 Jan 14. J Bacteriol. 2011. PMID: 21239589 Free PMC article.
-
myo-Inositol transport by Salmonella enterica serovar Typhimurium.Microbiology (Reading). 2010 Jan;156(Pt 1):128-138. doi: 10.1099/mic.0.032250-0. Epub 2009 Oct 15. Microbiology (Reading). 2010. PMID: 19833776
-
Hysteresis in myo-inositol utilization by Salmonella Typhimurium.Microbiologyopen. 2017 Apr;6(2):e00431. doi: 10.1002/mbo3.431. Epub 2016 Dec 27. Microbiologyopen. 2017. PMID: 28027601 Free PMC article.
-
Evolutionary Stability of Salmonella Competition with the Gut Microbiota: How the Environment Fosters Heterogeneity in Exploitative and Interference Competition.J Mol Biol. 2019 Nov 22;431(23):4732-4748. doi: 10.1016/j.jmb.2019.06.027. Epub 2019 Jun 28. J Mol Biol. 2019. PMID: 31260689 Review.
Cited by
-
Metabolism in the Niche: a Large-Scale Genome-Based Survey Reveals Inositol Utilization To Be Widespread among Soil, Commensal, and Pathogenic Bacteria.Microbiol Spectr. 2022 Aug 31;10(4):e0201322. doi: 10.1128/spectrum.02013-22. Epub 2022 Aug 4. Microbiol Spectr. 2022. PMID: 35924911 Free PMC article. Review.
-
The small RNA RssR regulates myo-inositol degradation by Salmonella enterica.Sci Rep. 2018 Dec 10;8(1):17739. doi: 10.1038/s41598-018-35784-8. Sci Rep. 2018. PMID: 30531898 Free PMC article.
-
Hierarchic regulation of a metabolic pathway: H-NS, CRP, and SsrB control myo-inositol utilization by Salmonella enterica.Microbiol Spectr. 2024 Jan 11;12(1):e0272423. doi: 10.1128/spectrum.02724-23. Epub 2023 Dec 14. Microbiol Spectr. 2024. PMID: 38095474 Free PMC article.
References
-
- Staib L. & Fuchs T. M. From food to cell: nutrient exploitation strategies of enteropathogens. Microbiology 160, 1020–1039 (2014). - PubMed
-
- Rothhardt J. E., Kröger C., Broadley S. P. & Fuchs T. M. The orphan regulator ReiD of Salmonella enterica is essential for myo-inositol utilization. Mol Microbiol 94, 700–712 (2014). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
