

Nonhomologous End-Joining with Minimal Sequence Loss Is Promoted by the Mre11-Rad50-Nbs1-Ctp1 Complex in Schizosaccharomyces pombe
- PMID: 28292918
- PMCID: PMC5419490
- DOI: 10.1534/genetics.117.200972
Nonhomologous End-Joining with Minimal Sequence Loss Is Promoted by the Mre11-Rad50-Nbs1-Ctp1 Complex in Schizosaccharomyces pombe
Abstract
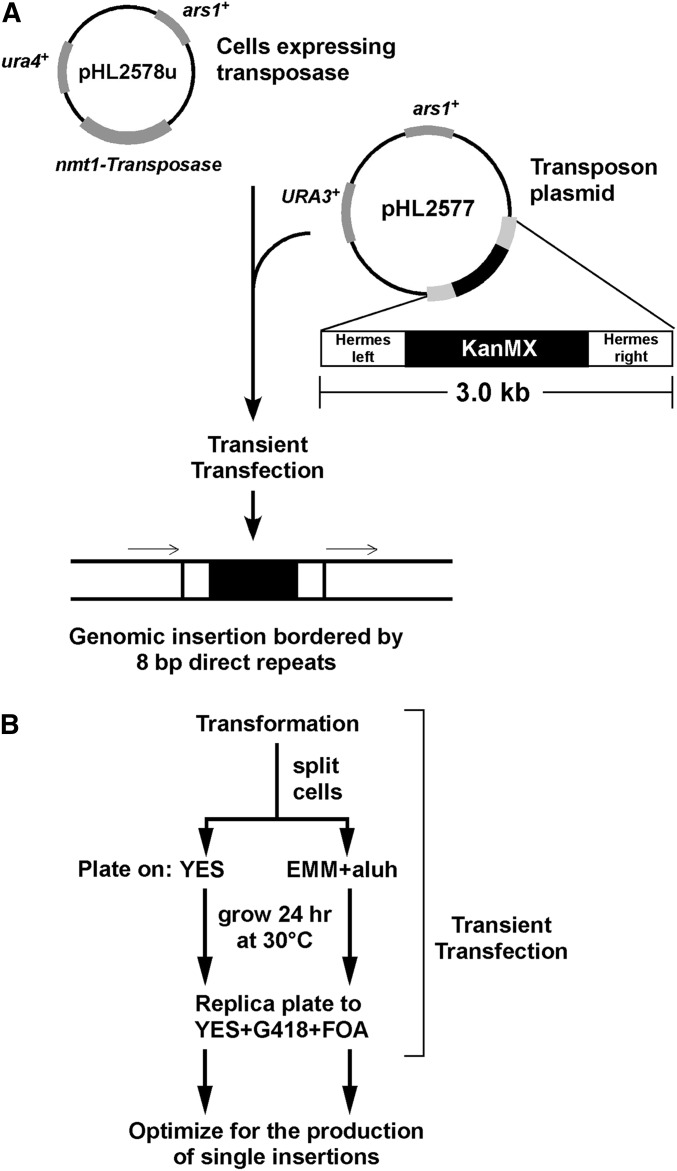
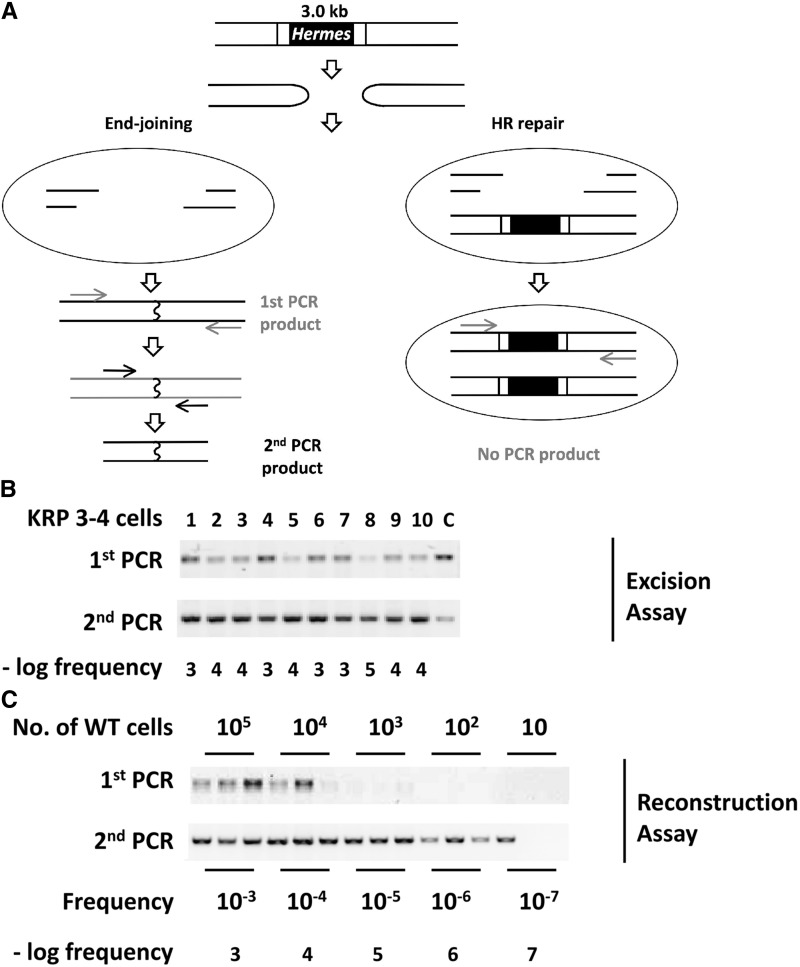
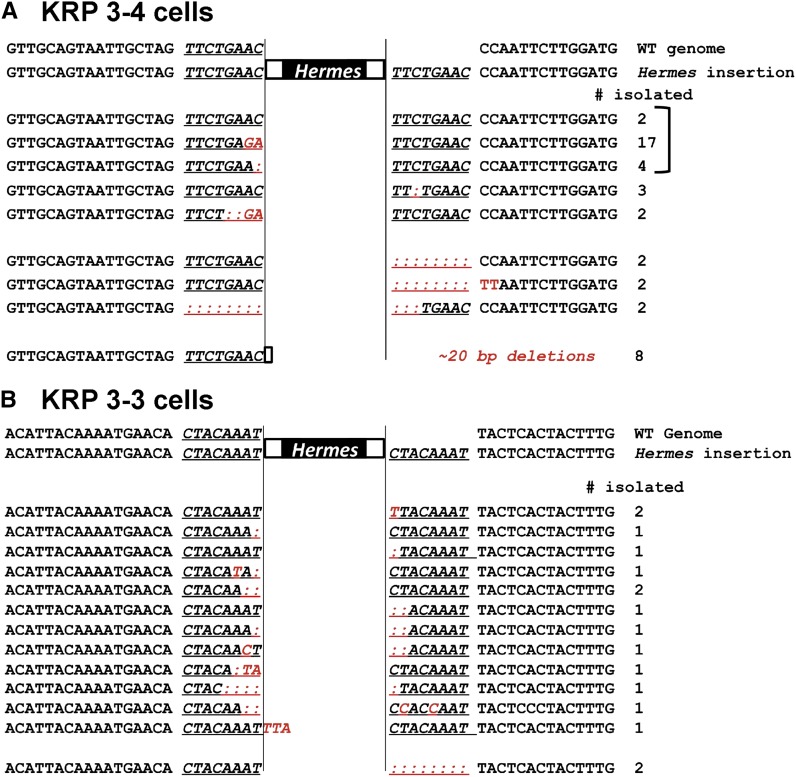
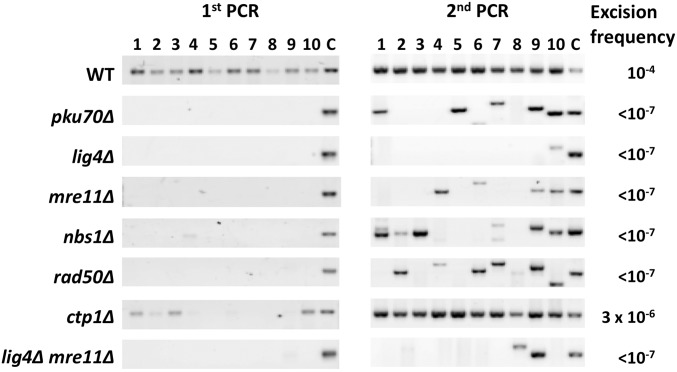
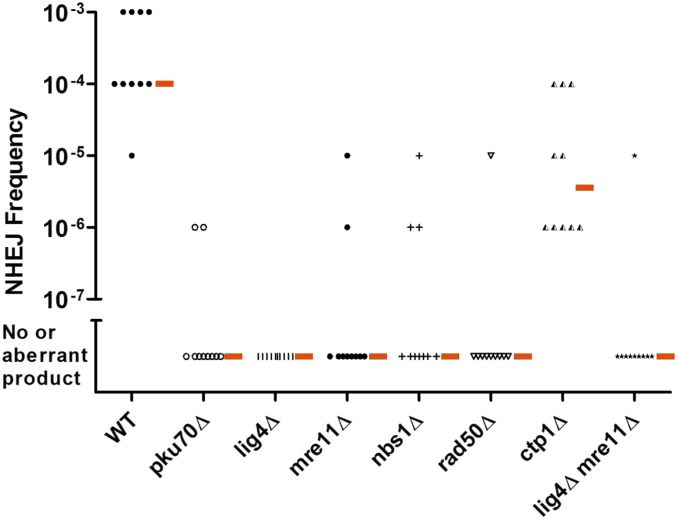
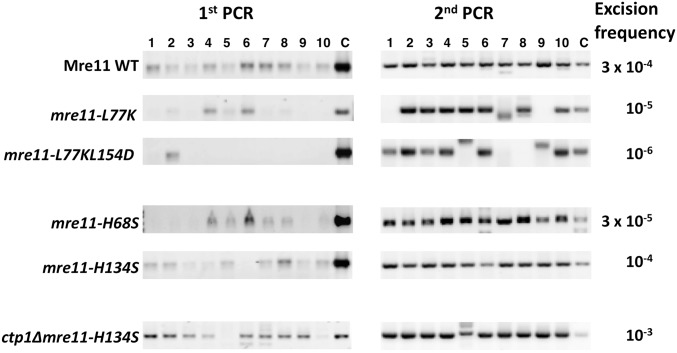
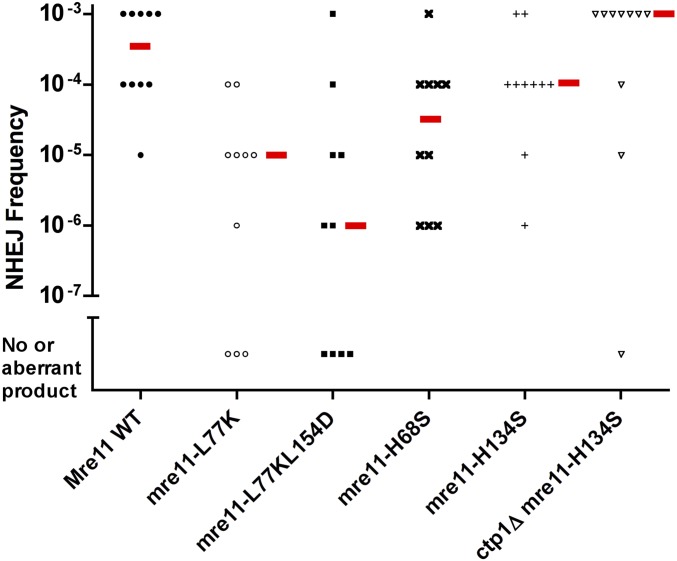
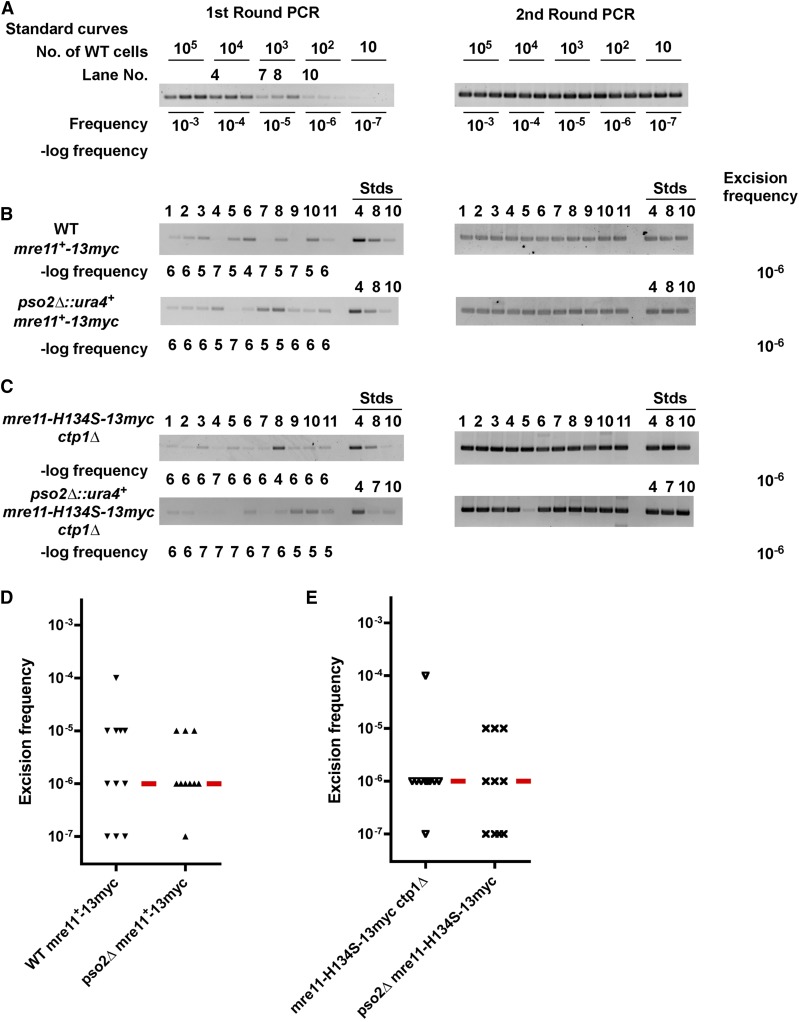
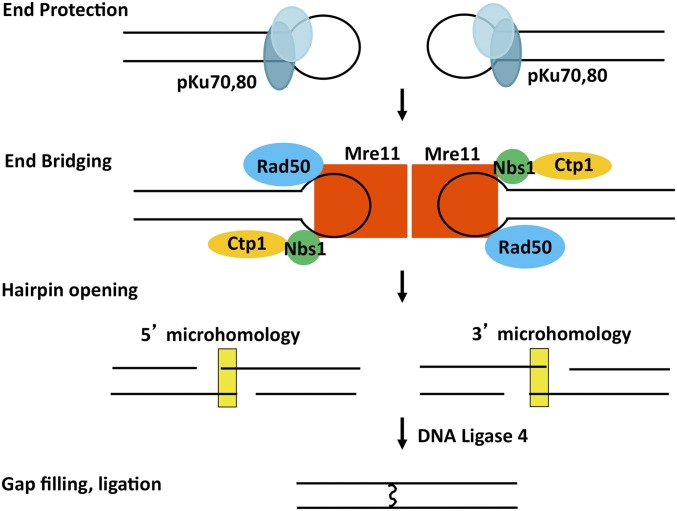
While the Mre11-Rad50-Nbs1 (MRN) complex has known roles in repair processes like homologous recombination and microhomology-mediated end-joining, its role in nonhomologous end-joining (NHEJ) is unclear as Saccharomyces cerevisiae, Schizosaccharomyces pombe, and mammals have different requirements for repairing cut DNA ends. Most double-strand breaks (DSBs) require nucleolytic processing prior to DNA ligation. Therefore, we studied repair using the Hermes transposon, whose excision leaves a DSB capped by hairpin ends similar to structures generated by palindromes and trinucleotide repeats. We generated single Hermes insertions using a novel S. pombe transient transfection system, and used Hermes excision to show a requirement for MRN in the NHEJ of nonligatable ends. NHEJ repair was indicated by the >1000-fold decrease in excision in cells lacking Ku or DNA ligase 4. Most repaired excision sites had <5 bp of sequence loss or mutation, characteristic for NHEJ and similar excision events in metazoans, and in contrast to the more extensive loss seen in S. cerevisiaeS. pombe NHEJ was reduced >1000-fold in cells lacking each MRN subunit, and loss of MRN-associated Ctp1 caused a 30-fold reduction. An Mre11 dimer is thought to hold DNA ends together for repair, and Mre11 dimerization domain mutations reduced repair 300-fold. In contrast, a mre11 mutant defective in endonucleolytic activity, the same mutant lacking Ctp1, or the triple mutant also lacking the putative hairpin nuclease Pso2 showed wild-type levels of repair. Thus, MRN may act to recruit the hairpin opening activity that allows subsequent repair.
Keywords: Hermes; MRN; MRX; Mre11-Rad50-Nbs1; Mre11-Rad50-Xrs2; NHEJ; hAT; hairpin; nonhomologous end-joining; transposon.
Copyright © 2017 by the Genetics Society of America.
Figures
References
-
- Arosio D., Cui S., Ortega C., Chovanec M., Di Marco S., et al. , 2002. Studies on the mode of Ku interaction with DNA. J. Biol. Chem. 277: 9741–9748. - PubMed
-
- Bähler J., Wood V., 2004. The genome and beyond, pp. 13–25 in The Molecular Biology of Schizosaccharomyces Pombe, edited by Egel R. Springer-Verlag, Berlin.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
