

Importance of the green color, absorption gradient, and spectral absorption of chloroplasts for the radiative energy balance of leaves
- PMID: 28293810
- PMCID: PMC5897488
- DOI: 10.1007/s10265-017-0910-z
Importance of the green color, absorption gradient, and spectral absorption of chloroplasts for the radiative energy balance of leaves
Erratum in
-
Correction to: Importance of the green color, absorption gradient, and spectral absorption of chloroplasts for the radiative energy balance of leaves.J Plant Res. 2018 May;131(3):569. doi: 10.1007/s10265-018-1014-0. J Plant Res. 2018. PMID: 29468324 Free PMC article.
Abstract
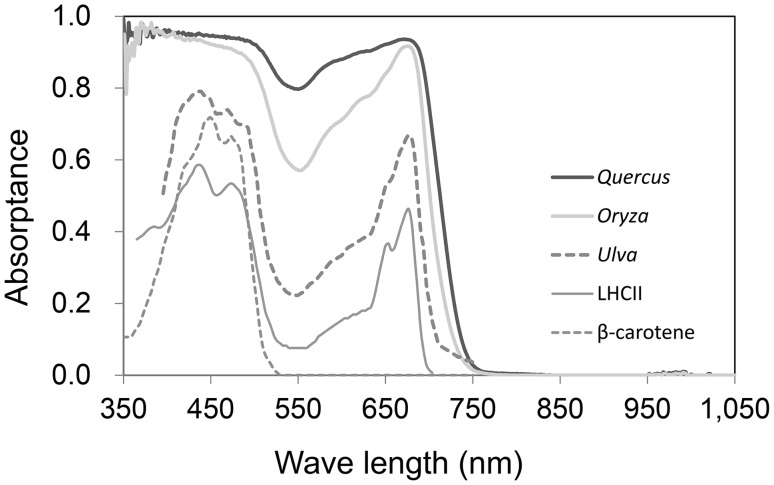
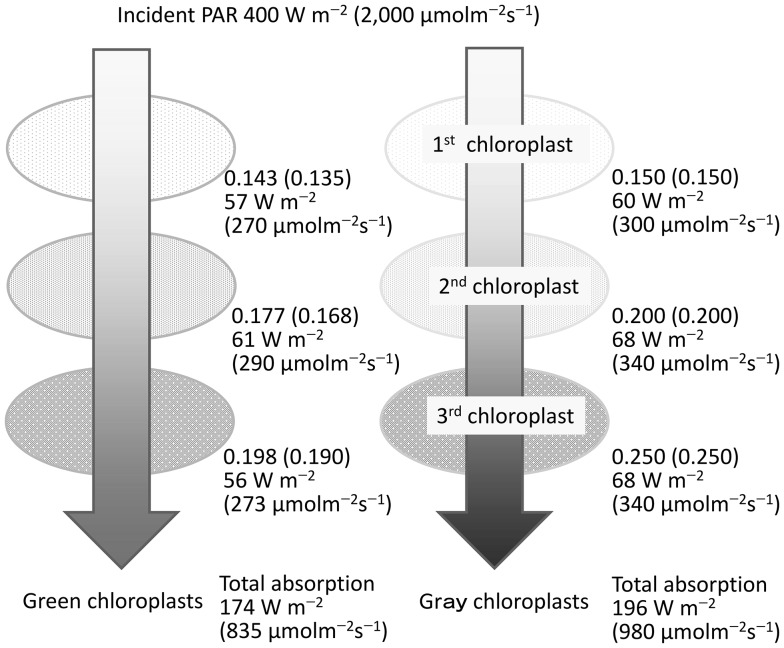
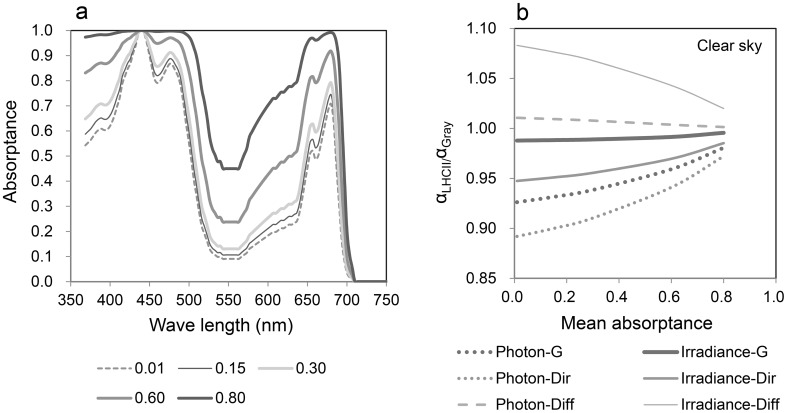
Terrestrial green plants absorb photosynthetically active radiation (PAR; 400-700 nm) but do not absorb photons evenly across the PAR waveband. The spectral absorbance of photosystems and chloroplasts is lowest for green light, which occurs within the highest irradiance waveband of direct solar radiation. We demonstrate a close relationship between this phenomenon and the safe and efficient utilization of direct solar radiation in simple biophysiological models. The effects of spectral absorptance on the photon and irradiance absorption processes are evaluated using the spectra of direct and diffuse solar radiation. The radiation absorption of a leaf arises as a consequence of the absorption of chloroplasts. The photon absorption of chloroplasts is strongly dependent on the distribution of pigment concentrations and their absorbance spectra. While chloroplast movements in response to light are important mechanisms controlling PAR absorption, they are not effective for green light because chloroplasts have the lowest spectral absorptance in the waveband. With the development of palisade tissue, the incident photons per total palisade cell surface area and the absorbed photons per chloroplast decrease. The spectral absorbance of carotenoids is effective in eliminating shortwave PAR (<520 nm), which contains much of the surplus energy that is not used for photosynthesis and is dissipated as heat. The PAR absorptance of a whole leaf shows no substantial difference based on the spectra of direct or diffuse solar radiation. However, most of the near infrared radiation is unabsorbed and heat stress is greatly reduced. The incident solar radiation is too strong to be utilized for photosynthesis under the current CO2 concentration in the terrestrial environment. Therefore, the photon absorption of a whole leaf is efficiently regulated by photosynthetic pigments with low spectral absorptance in the highest irradiance waveband and through a combination of pigment density distribution and leaf anatomical structures.
Keywords: Absorption spectra; Carotenoids; Chloroplast movement; Direct radiation; Palisade tissue; Photosystem.
Figures







References
-
- Akitsu T, Kume A, Hirose Y, Ijima O, Nasahara KN. On the stability of radiometric ratios of photosynthetically active radiation to global solar radiation in Tsukuba, Japan. Agric For Meteorol. 2015;209–210:59–68. doi: 10.1016/j.agrformet.2015.04.026. - DOI
-
- Brodersen CR, Vogelmann TC. Do changes in light direction affect absorption profile in leaves? Funct Plant Biol. 2010;37:403–412. doi: 10.1071/FP09262. - DOI
-
- Brodersen CR, Vogelmann TC, Williams WE, Gorton HL. A new paradigm in leaf-level photosynthesis: direct and diffuse lights are not equal. Plant Cell Environ. 2008;31:159–164. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
