

Nitric oxide-enhanced Shiga toxin production was regulated by Fur and RecA in enterohemorrhagic Escherichia coli O157
- PMID: 28294553
- PMCID: PMC5552940
- DOI: 10.1002/mbo3.461
Nitric oxide-enhanced Shiga toxin production was regulated by Fur and RecA in enterohemorrhagic Escherichia coli O157
Abstract
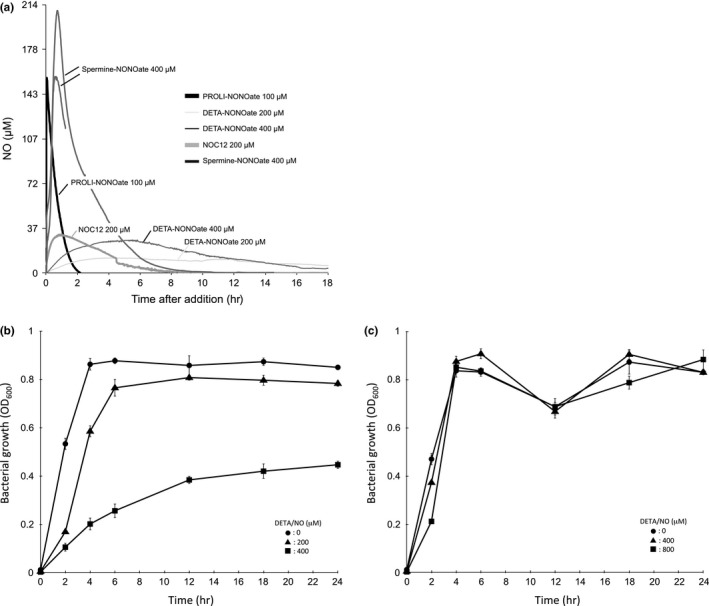
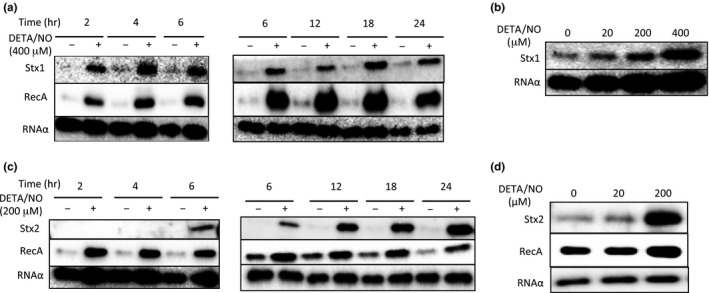
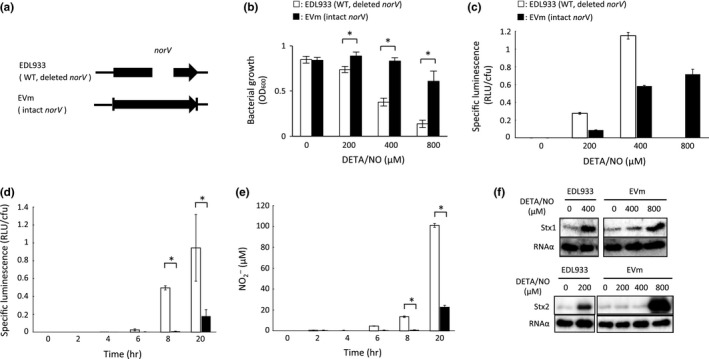
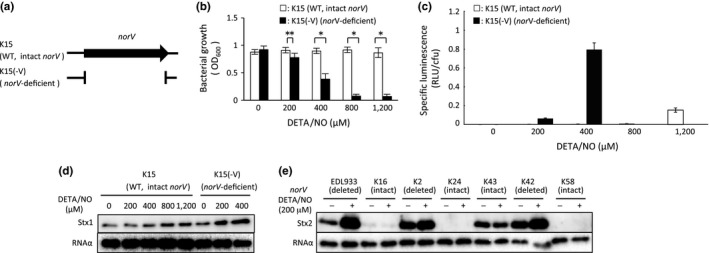
Enterohemorrhagic Escherichia coli (EHEC) produces Shiga toxin 1 (Stx1) and Shiga toxin 2 (Stx2). Nitric oxide (NO), which acts as an antimicrobial defense molecule, was found to enhance the production of Stx1 and Stx2 in EHEC under anaerobic conditions. Although EHEC O157 has two types of anaerobic NO reductase genes, an intact norV and a deleted norV, in the deleted norV-type EHEC, a high concentration of NO (12-29 μmol/L, maximum steady-state concentration) is required for enhanced Stx1 production and a low concentration of NO (~12 μmol/L, maximum steady-state concentration) is sufficient for enhanced Stx2 production under anaerobic conditions. These results suggested that different concentration thresholds of NO elicit a discrete set of Stx1 and Stx2 production pathways. Moreover, the enhancement of Shiga toxin production in the intact norV-type EHEC required treatment with a higher concentration of NO than was required for enhancement of Shiga toxin production in the deleted norV-type EHEC, suggesting that the specific NorV type plays an important role in the level of enhancement of Shiga toxin production in response to NO. Finally, Fur derepression and RecA activation in EHEC were shown to participate in the NO-enhanced Stx1 and Stx2 production, respectively.
Keywords: Fur; NO reductase; RecA; Shiga toxin; enterohemorrhagic Escherichia coli; nitric oxide.
© 2017 The Authors. MicrobiologyOpen published by John Wiley & Sons Ltd.
Figures
References
-
- Bogdan, C. (2001). Nitric oxide and the immune response. Nature Immunology, 2, 907–916. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
