

Let-7 and MicroRNA-148 Regulate Parathyroid Hormone Levels in Secondary Hyperparathyroidism
- PMID: 28298326
- PMCID: PMC5533223
- DOI: 10.1681/ASN.2016050585
Let-7 and MicroRNA-148 Regulate Parathyroid Hormone Levels in Secondary Hyperparathyroidism
Abstract
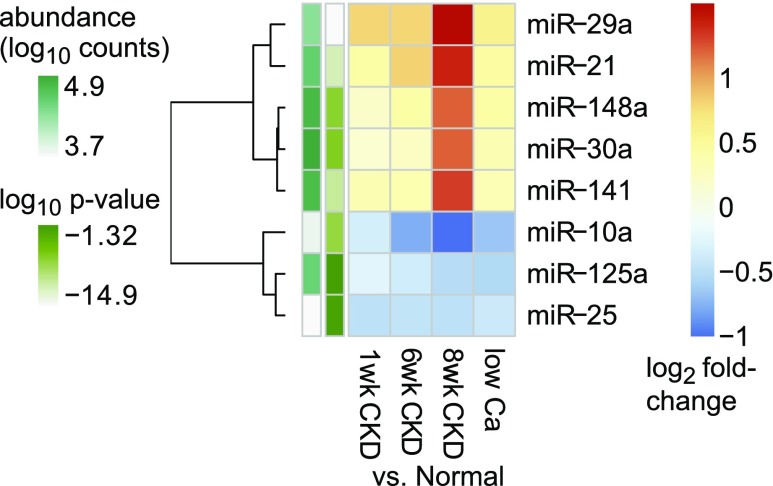
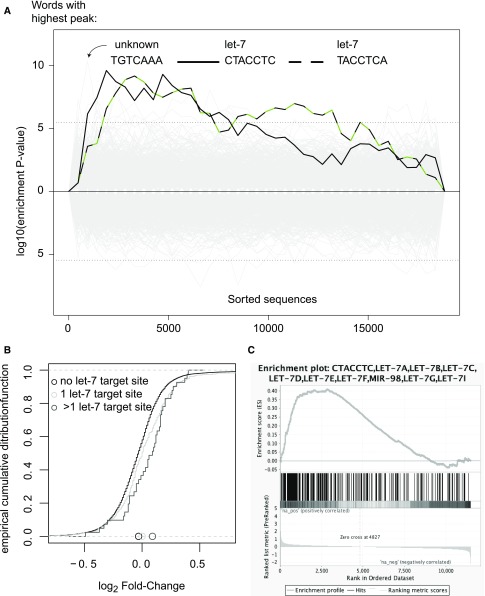
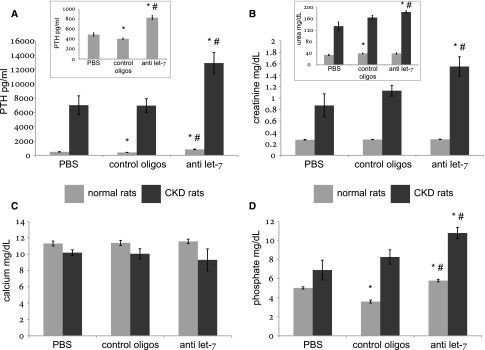
Secondary hyperparathyroidism commonly complicates CKD and associates with morbidity and mortality. We profiled microRNA (miRNA) in parathyroid glands from experimental hyperparathyroidism models and patients receiving dialysis and studied the function of specific miRNAs. miRNA deep-sequencing showed that human and rodent parathyroids share similar profiles. Parathyroids from uremic and normal rats segregated on the basis of their miRNA expression profiles, and a similar finding was observed in humans. We identified parathyroid miRNAs that were dysregulated in experimental hyperparathyroidism, including miR-29, miR-21, miR-148, miR-30, and miR-141 (upregulated); and miR-10, miR-125, and miR-25 (downregulated). Inhibition of the abundant let-7 family increased parathyroid hormone (PTH) secretion in normal and uremic rats, as well as in mouse parathyroid organ cultures. Conversely, inhibition of the upregulated miR-148 family prevented the increase in serum PTH level in uremic rats and decreased levels of secreted PTH in parathyroid cultures. The evolutionary conservation of abundant miRNAs in normal parathyroid glands and the regulation of these miRNAs in secondary hyperparathyroidism indicates their importance for parathyroid function and the development of hyperparathyroidism. Specifically, let-7 and miR-148 antagonism modified PTH secretion in vivo and in vitro, implying roles for these specific miRNAs. These findings may be utilized for therapeutic interventions aimed at altering PTH expression in diseases such as osteoporosis and secondary hyperparathyroidism.
Keywords: chronic kidney disease; gene expression; hyperparathyroidism; microRNA; mineral metabolism.
Copyright © 2017 by the American Society of Nephrology.
Figures





References
-
- Farazi TA, Horlings HM, Ten Hoeve JJ, Mihailovic A, Halfwerk H, Morozov P, Brown M, Hafner M, Reyal F, van Kouwenhove M, Kreike B, Sie D, Hovestadt V, Wessels LFA, van de Vijver MJ, Tuschl T: MicroRNA sequence and expression analysis in breast tumors by deep sequencing. Cancer Res 71: 4443–4453, 2011 - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
