

Network reorganization and breakdown of an ant-plant protection mutualism with elevation
- PMID: 28298349
- PMCID: PMC5360921
- DOI: 10.1098/rspb.2016.2564
Network reorganization and breakdown of an ant-plant protection mutualism with elevation
Abstract
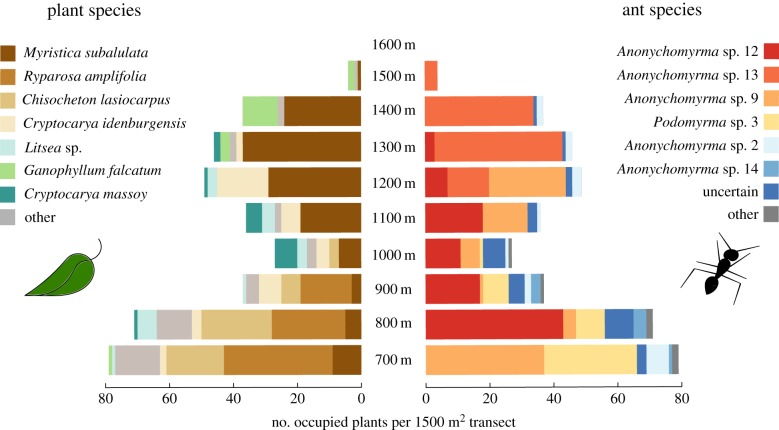
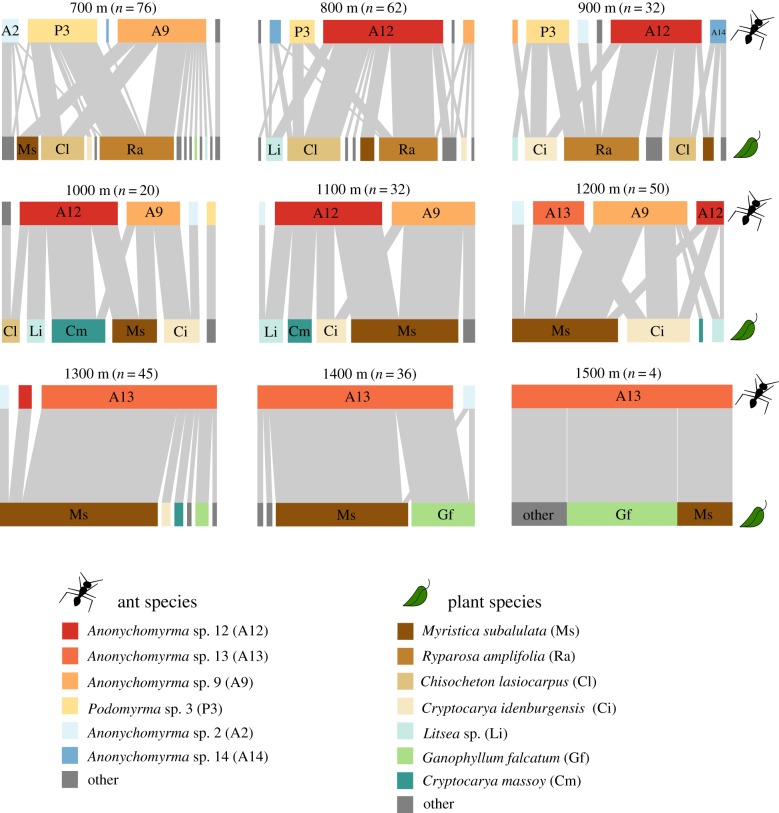
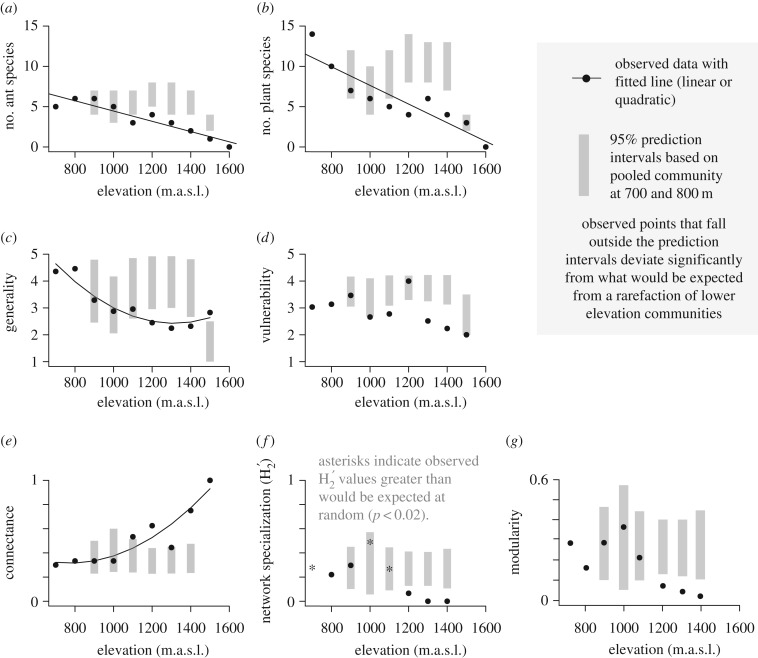
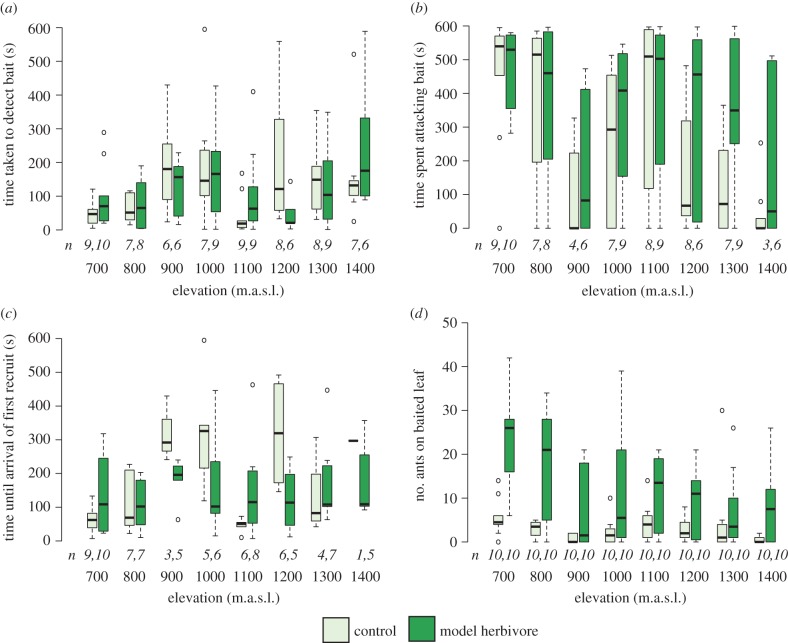
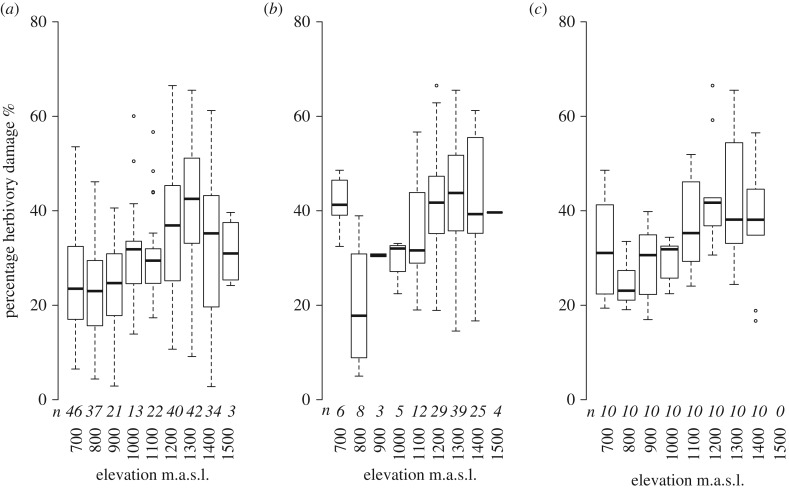
Both the abiotic environment and the composition of animal and plant communities change with elevation. For mutualistic species, these changes are expected to result in altered partner availability, and shifts in context-dependent benefits for partners. To test these predictions, we assessed the network structure of terrestrial ant-plant mutualists and how the benefits to plants of ant inhabitation changed with elevation in tropical forest in Papua New Guinea. At higher elevations, ant-plants were rarer, species richness of both ants and plants decreased, and the average ant or plant species interacted with fewer partners. However, networks became increasingly connected and less specialized, more than could be accounted for by reductions in ant-plant abundance. On the most common ant-plant, ants recruited less and spent less time attacking a surrogate herbivore at higher elevations, and herbivory damage increased. These changes were driven by turnover of ant species rather than by within-species shifts in protective behaviour. We speculate that reduced partner availability at higher elevations results in less specialized networks, while lower temperatures mean that even for ant-inhabited plants, benefits are reduced. Under increased abiotic stress, mutualistic networks can break down, owing to a combination of lower population sizes, and a reduction in context-dependent mutualistic benefits.
Keywords: altitudinal gradients; biotic defence; global change; herbivory; myrmecophyte; network specialization.
© 2017 The Author(s).
Conflict of interest statement
The authors have no competing interests.
Figures

Similar articles
-
Effects of dam-induced landscape fragmentation on amazonian ant-plant mutualistic networks.Conserv Biol. 2013 Aug;27(4):763-73. doi: 10.1111/cobi.12045. Epub 2013 Apr 2. Conserv Biol. 2013. PMID: 23551525
-
Effects of chronic anthropogenic disturbance and rainfall on the specialization of ant-plant mutualistic networks in the Caatinga, a Brazilian dry forest.J Anim Ecol. 2018 Jul;87(4):1022-1033. doi: 10.1111/1365-2656.12820. Epub 2018 Apr 6. J Anim Ecol. 2018. PMID: 29504629
-
Urbanization drives partner switching and loss of mutualism in an ant-plant symbiosis.Ecology. 2024 Nov;105(11):e4449. doi: 10.1002/ecy.4449. Epub 2024 Oct 14. Ecology. 2024. PMID: 39400307
-
Current issues in the evolutionary ecology of ant-plant symbioses.New Phytol. 2014 May;202(3):749-764. doi: 10.1111/nph.12690. Epub 2014 Jan 21. New Phytol. 2014. PMID: 24444030 Review.
-
Ant predation on herbivores through a multitrophic lens: how effects of ants on plant herbivore defense and natural enemies vary along temperature gradients.Curr Opin Insect Sci. 2016 Apr;14:73-80. doi: 10.1016/j.cois.2016.02.001. Epub 2016 Feb 8. Curr Opin Insect Sci. 2016. PMID: 27436650 Review.
Cited by
-
Phylogenetic structure of moth communities (Geometridae, Lepidoptera) along a complete rainforest elevational gradient in Papua New Guinea.PLoS One. 2024 Aug 12;19(8):e0308698. doi: 10.1371/journal.pone.0308698. eCollection 2024. PLoS One. 2024. PMID: 39133743 Free PMC article.
-
Refuges from fire maintain pollinator-plant interaction networks.Ecol Evol. 2019 Apr 30;9(10):5777-5786. doi: 10.1002/ece3.5161. eCollection 2019 May. Ecol Evol. 2019. PMID: 31160998 Free PMC article.
-
The turnover of plant-frugivore interactions along plant range expansion: consequences for natural colonization processes.Proc Biol Sci. 2023 May 31;290(1999):20222547. doi: 10.1098/rspb.2022.2547. Epub 2023 May 24. Proc Biol Sci. 2023. PMID: 37221844 Free PMC article.
-
Highly modular pattern in ant-plant interactions involving specialized and non-specialized myrmecophytes.Naturwissenschaften. 2018 Jun 27;105(7-8):43. doi: 10.1007/s00114-018-1570-0. Naturwissenschaften. 2018. PMID: 29951968
-
The interactions of ants with their biotic environment.Proc Biol Sci. 2017 Mar 15;284(1850):20170013. doi: 10.1098/rspb.2017.0013. Proc Biol Sci. 2017. PMID: 28298352 Free PMC article. Review.
References
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
