

Scavenger Receptor MARCO Orchestrates Early Defenses and Contributes to Fungal Containment during Cryptococcal Infection
- PMID: 28298522
- PMCID: PMC5423401
- DOI: 10.4049/jimmunol.1700057
Scavenger Receptor MARCO Orchestrates Early Defenses and Contributes to Fungal Containment during Cryptococcal Infection
Abstract
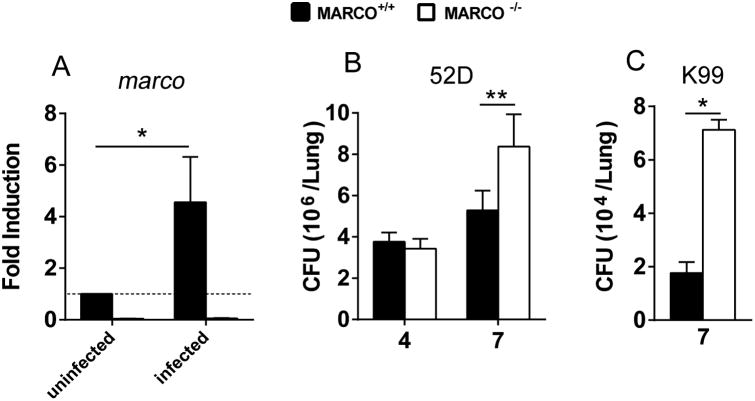
The scavenger receptor macrophage receptor with collagenous structure (MARCO) promotes protective innate immunity against bacterial and parasitic infections; however, its role in host immunity against fungal pathogens, including the major human opportunistic fungal pathogen Cryptococcus neoformans, remains unknown. Using a mouse model of C. neoformans infection, we demonstrated that MARCO deficiency leads to impaired fungal control during the afferent phase of cryptococcal infection. Diminished fungal containment in MARCO-/- mice was accompanied by impaired recruitment of Ly6Chigh monocytes and monocyte-derived dendritic cells (moDC) and lower moDC costimulatory maturation. The reduced recruitment and activation of mononuclear phagocytes in MARCO-/- mice was linked to diminished early expression of IFN-γ along with profound suppression of CCL2 and CCL7 chemokines, providing evidence for roles of MARCO in activation of the CCR2 axis during C. neoformans infection. Lastly, we found that MARCO was involved in C. neoformans phagocytosis by resident pulmonary macrophages and DC. We conclude that MARCO facilitates early interactions between C. neoformans and lung-resident cells and promotes the production of CCR2 ligands. In turn, this contributes to a more robust recruitment and activation of moDC that opposes rapid fungal expansion during the afferent phase of cryptococcal infection.
Copyright © 2017 by The American Association of Immunologists, Inc.
Figures








References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
