

African Swine Fever Virus NP868R Capping Enzyme Promotes Reovirus Rescue during Reverse Genetics by Promoting Reovirus Protein Expression, Virion Assembly, and RNA Incorporation into Infectious Virions
- PMID: 28298603
- PMCID: PMC5432856
- DOI: 10.1128/JVI.02416-16
African Swine Fever Virus NP868R Capping Enzyme Promotes Reovirus Rescue during Reverse Genetics by Promoting Reovirus Protein Expression, Virion Assembly, and RNA Incorporation into Infectious Virions
Abstract
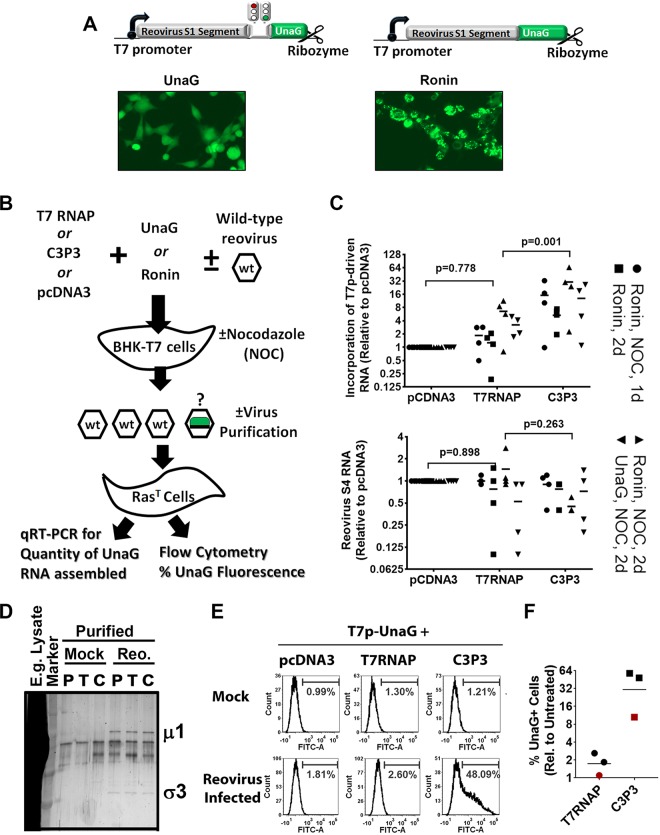
Reoviruses, like many eukaryotic viruses, contain an inverted 7-methylguanosine (m7G) cap linked to the 5' nucleotide of mRNA. The traditional functions of capping are to promote mRNA stability, protein translation, and concealment from cellular proteins that recognize foreign RNA. To address the role of mRNA capping during reovirus replication, we assessed the benefits of adding the African swine fever virus NP868R capping enzyme during reovirus rescue. C3P3, a fusion protein containing T7 RNA polymerase and NP868R, was found to increase protein expression 5- to 10-fold compared to T7 RNA polymerase alone while enhancing reovirus rescue from the current reverse genetics system by 100-fold. Surprisingly, RNA stability was not increased by C3P3, suggesting a direct effect on protein translation. A time course analysis revealed that C3P3 increased protein synthesis within the first 2 days of a reverse genetics transfection. This analysis also revealed that C3P3 enhanced processing of outer capsid μ1 protein to μ1C, a previously described hallmark of reovirus assembly. Finally, to determine the rate of infectious-RNA incorporation into new virions, we developed a new recombinant reovirus S1 gene that expressed the fluorescent protein UnaG. Following transfection of cells with UnaG and infection with wild-type virus, passage of UnaG through progeny was significantly enhanced by C3P3. These data suggest that capping provides nontraditional functions to reovirus, such as promoting assembly and infectious-RNA incorporation.IMPORTANCE Our findings expand our understanding of how viruses utilize capping, suggesting that capping provides nontraditional functions to reovirus, such as promoting assembly and infectious-RNA incorporation, in addition to enhancing protein translation. Beyond providing mechanistic insight into reovirus replication, our findings also show that reovirus reverse genetics rescue is enhanced 100-fold by the NP868R capping enzyme. Since reovirus shows promise as a cancer therapy, efficient reovirus reverse genetics rescue will accelerate production of recombinant reoviruses as candidates to enhance therapeutic potency. NP868R-assisted reovirus rescue will also expedite production of recombinant reovirus for mechanistic insights into reovirus protein function and structure.
Keywords: C3P3; NP868R; capping; reovirus; reverse genetics.
Copyright © 2017 American Society for Microbiology.
Figures





Similar articles
-
Generation of Recombinant Rotavirus Expressing NSP3-UnaG Fusion Protein by a Simplified Reverse Genetics System.J Virol. 2019 Nov 26;93(24):e01616-19. doi: 10.1128/JVI.01616-19. Print 2019 Dec 15. J Virol. 2019. PMID: 31597761 Free PMC article.
-
Reverse genetics for mammalian reovirus.Methods. 2011 Oct;55(2):109-13. doi: 10.1016/j.ymeth.2011.07.002. Epub 2011 Jul 21. Methods. 2011. PMID: 21798351 Free PMC article.
-
An Optimized Reverse Genetics System Suitable for Efficient Recovery of Simian, Human, and Murine-Like Rotaviruses.J Virol. 2020 Aug 31;94(18):e01294-20. doi: 10.1128/JVI.01294-20. Print 2020 Aug 31. J Virol. 2020. PMID: 32759316 Free PMC article.
-
Control of Capsid Transformations during Reovirus Entry.Viruses. 2021 Jan 21;13(2):153. doi: 10.3390/v13020153. Viruses. 2021. PMID: 33494426 Free PMC article. Review.
-
[Orthoreoviruses].Uirusu. 2014;64(2):191-202. doi: 10.2222/jsv.64.191. Uirusu. 2014. PMID: 26437841 Review. Japanese.
Cited by
-
Expression of Separate Heterologous Proteins from the Rotavirus NSP3 Genome Segment Using a Translational 2A Stop-Restart Element.J Virol. 2020 Aug 31;94(18):e00959-20. doi: 10.1128/JVI.00959-20. Print 2020 Aug 31. J Virol. 2020. PMID: 32611753 Free PMC article.
-
Single Amino Acid Differences between Closely Related Reovirus T3D Lab Strains Alter Oncolytic Potency In Vitro and In Vivo.J Virol. 2020 Jan 31;94(4):e01688-19. doi: 10.1128/JVI.01688-19. Print 2020 Jan 31. J Virol. 2020. PMID: 31748391 Free PMC article.
-
Development of an entirely plasmid-based reverse genetics system for 12-segmented double-stranded RNA viruses.Proc Natl Acad Sci U S A. 2021 Oct 19;118(42):e2105334118. doi: 10.1073/pnas.2105334118. Proc Natl Acad Sci U S A. 2021. PMID: 34635593 Free PMC article.
-
Mature Rotavirus Particles Contain Equivalent Amounts of 7meGpppG-Capped and Noncapped Viral Positive-Sense RNAs.J Virol. 2022 Sep 14;96(17):e0115122. doi: 10.1128/jvi.01151-22. Epub 2022 Aug 24. J Virol. 2022. PMID: 36000838 Free PMC article.
-
C3P3-G1: first generation of a eukaryotic artificial cytoplasmic expression system.Nucleic Acids Res. 2019 Mar 18;47(5):2681-2698. doi: 10.1093/nar/gkz069. Nucleic Acids Res. 2019. PMID: 30726994 Free PMC article.
References
-
- Picard-Jean F, Tremblay-Letourneau M, Serra E, Dimech C, Schulz H, Anselin M, Dutilly V, Bisaillon M. 2013. RNA 5′-end maturation: a crucial step in the replication of viral genomes. In Romanowski V. (ed), Current issues in molecular virology: viral genetics and biotechnological applications. In Tech, Rijeka, Croatia. doi: 10.5772/56166. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
