

Frequency tuning and directional sensitivity of tympanal vibrations in the field cricket Gryllus bimaculatus
- PMID: 28298611
- PMCID: PMC5378147
- DOI: 10.1098/rsif.2017.0035
Frequency tuning and directional sensitivity of tympanal vibrations in the field cricket Gryllus bimaculatus
Abstract
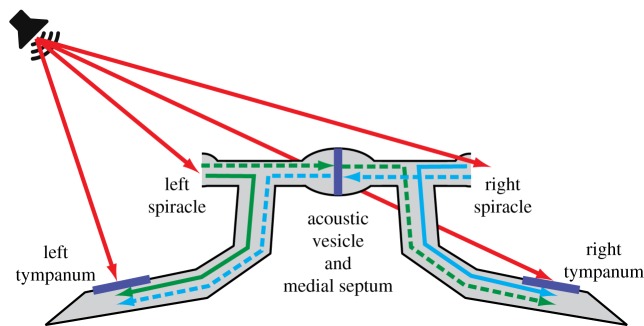
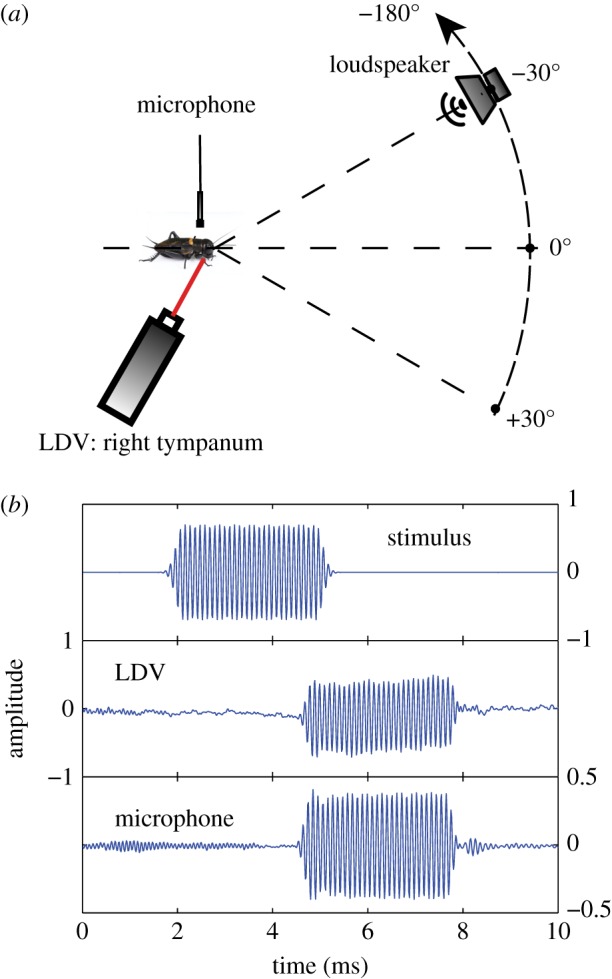
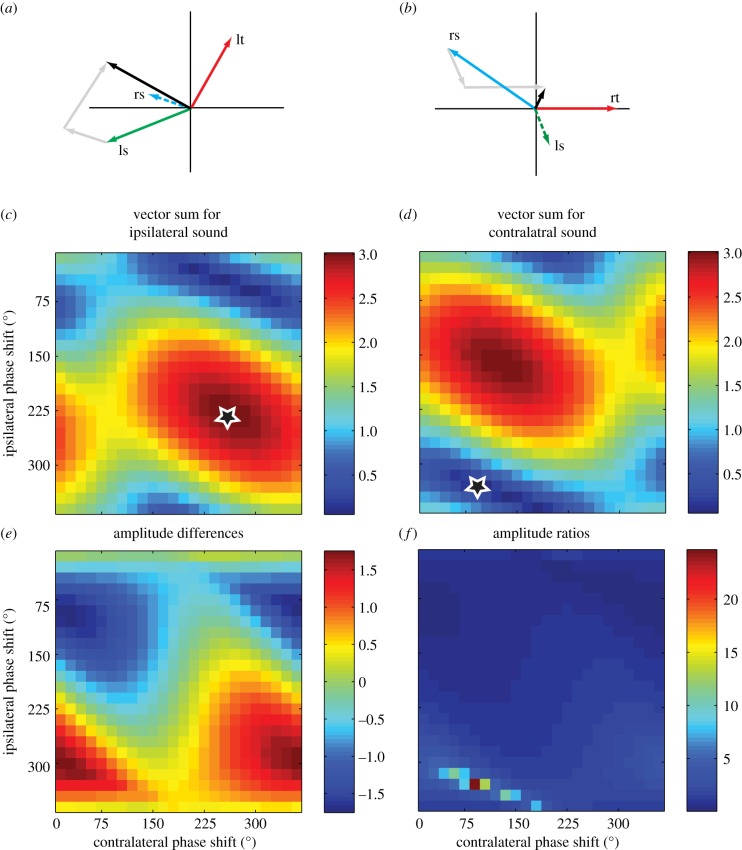
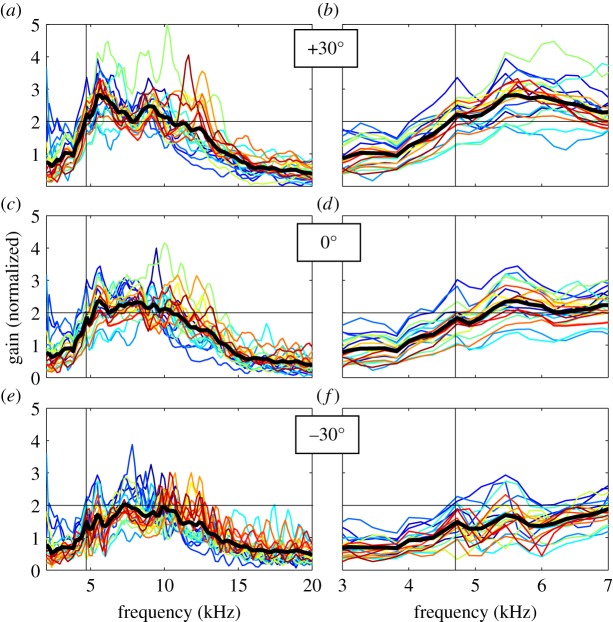
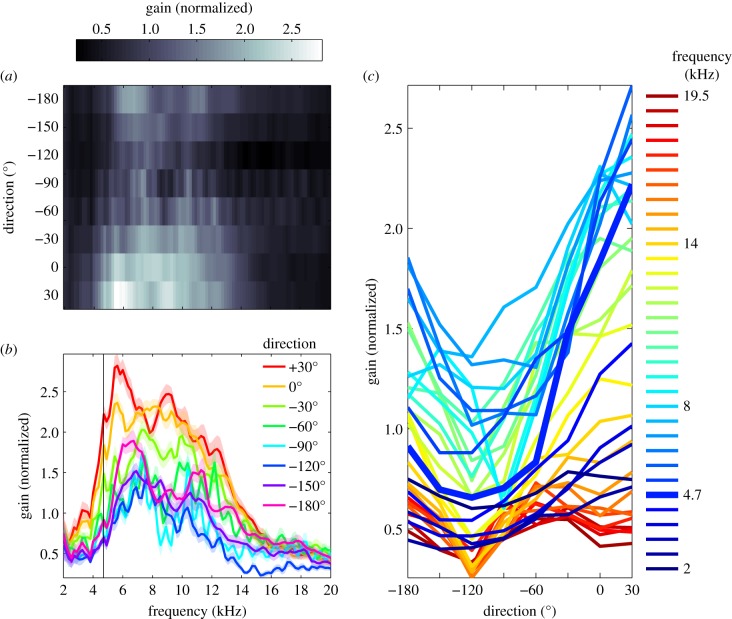
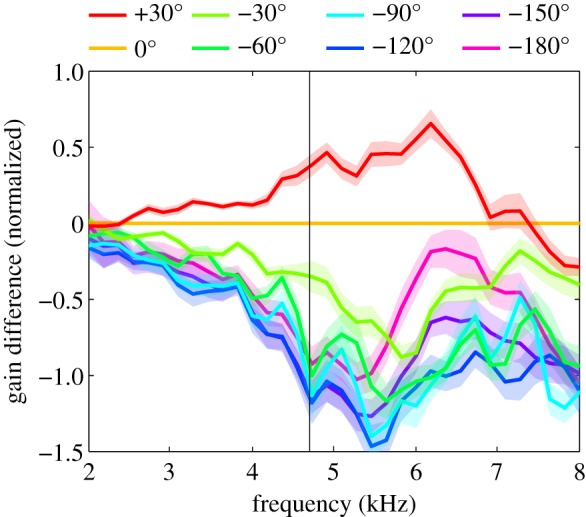
Female field crickets use phonotaxis to locate males by their calling song. Male song production and female behavioural sensitivity form a pair of matched frequency filters, which in Gryllus bimaculatus are tuned to a frequency of about 4.7 kHz. Directional sensitivity is supported by an elaborate system of acoustic tracheae, which make the ears function as pressure difference receivers. As a result, phase differences between left and right sound inputs are transformed into vibration amplitude differences. Here we critically tested the hypothesis that acoustic properties of internal transmissions play a major role in tuning directional sensitivity to the calling song frequency, by measuring tympanal vibrations as a function of sound direction and frequency. Rather than sharp frequency tuning of directional sensitivity corresponding to the calling song, we found broad frequency tuning, with optima shifted to higher frequencies. These findings agree with predictions from a vector summation model for combining external and internal sounds. We show that the model provides robust directional sensitivity that is, however, broadly tuned with an optimum well above the calling song frequency. We therefore advocate that additional filtering, e.g. at a higher (neuronal) level, significantly contributes to frequency tuning of directional sensitivity.
Keywords: directional hearing; laser Doppler vibrometer; phase shift; phonotaxis; pressure difference receiver.
© 2017 The Author(s).
Figures
References
MeSH terms
Associated data
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials