

Functional Roles of the Interaction of APP and Lipoprotein Receptors
- PMID: 28298885
- PMCID: PMC5331069
- DOI: 10.3389/fnmol.2017.00054
Functional Roles of the Interaction of APP and Lipoprotein Receptors
Abstract
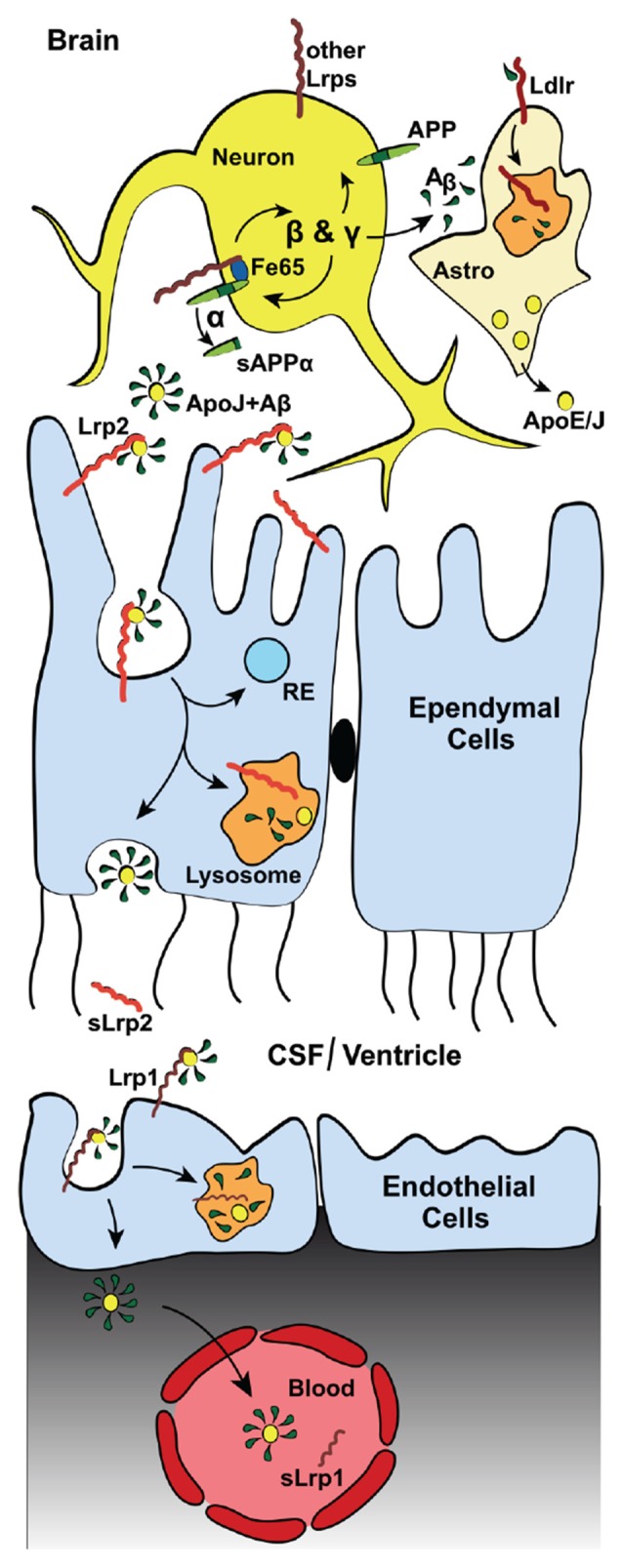
The biological fates of the key initiator of Alzheimer's disease (AD), the amyloid precursor protein (APP), and a family of lipoprotein receptors, the low-density lipoprotein (LDL) receptor-related proteins (LRPs) and their molecular roles in the neurodegenerative disease process are inseparably interwoven. Not only does APP bind tightly to the extracellular domains (ECDs) of several members of the LRP group, their intracellular portions are also connected through scaffolds like the one established by FE65 proteins and through interactions with adaptor proteins such as X11/Mint and Dab1. Moreover, the ECDs of APP and LRPs share common ligands, most notably Reelin, a regulator of neuronal migration during embryonic development and modulator of synaptic transmission in the adult brain, and Agrin, another signaling protein which is essential for the formation and maintenance of the neuromuscular junction (NMJ) and which likely also has critical, though at this time less well defined, roles for the regulation of central synapses. Furthermore, the major independent risk factors for AD, Apolipoprotein (Apo) E and ApoJ/Clusterin, are lipoprotein ligands for LRPs. Receptors and ligands mutually influence their intracellular trafficking and thereby the functions and abilities of neurons and the blood-brain-barrier to turn over and remove the pathological product of APP, the amyloid-β peptide. This article will review and summarize the molecular mechanisms that are shared by APP and LRPs and discuss their relative contributions to AD.
Keywords: APOE; LDL receptor gene family; LRP; amyloid beta; glutamate receptors; neuromuscular junction; synapse; trafficking.
Figures



References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous