

H3 ubiquitination by NEDD4 regulates H3 acetylation and tumorigenesis
- PMID: 28300060
- PMCID: PMC5357315
- DOI: 10.1038/ncomms14799
H3 ubiquitination by NEDD4 regulates H3 acetylation and tumorigenesis
Abstract
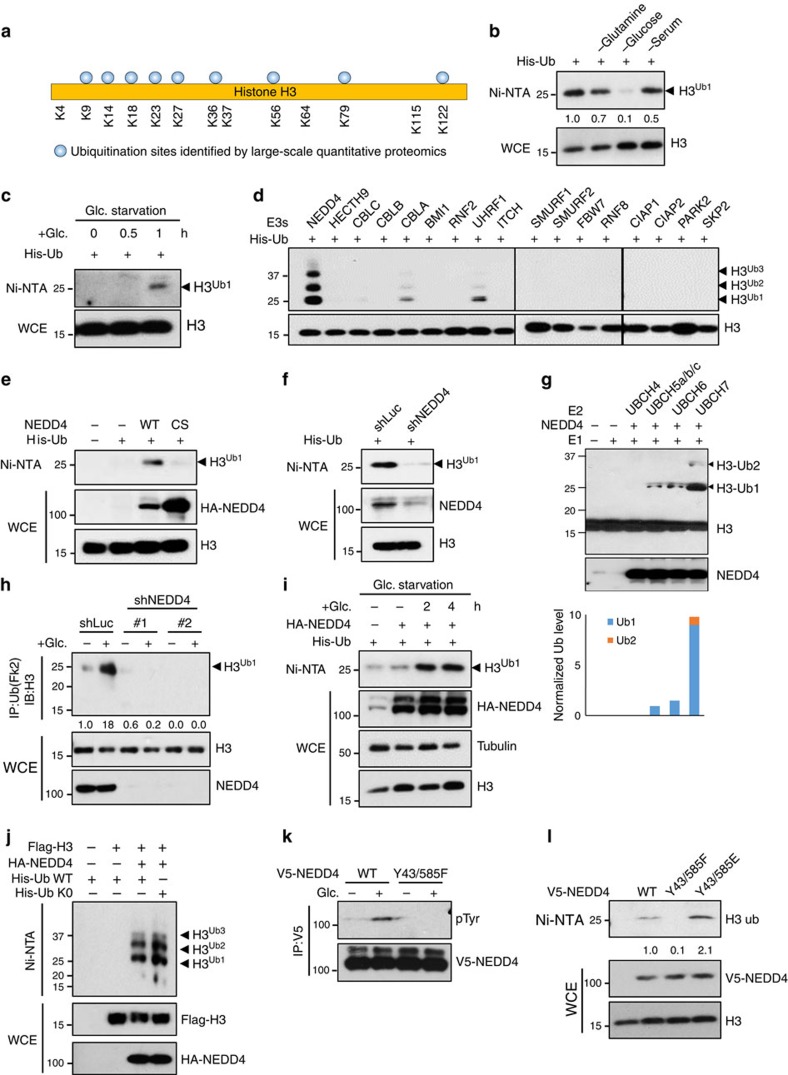
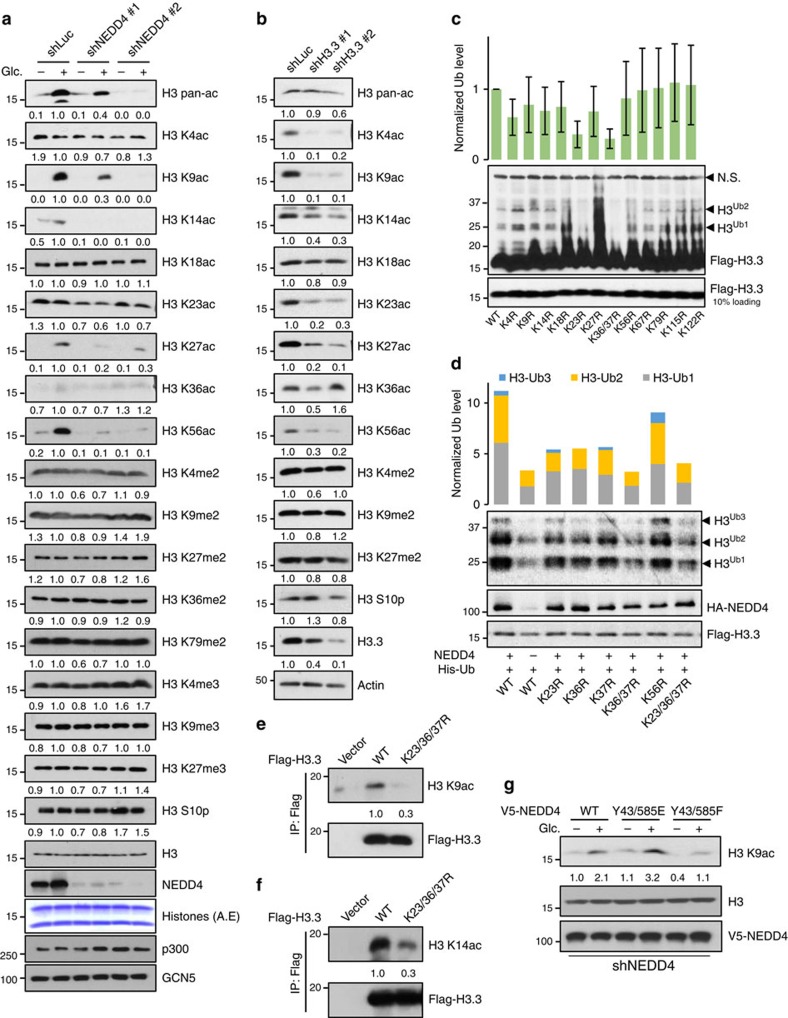
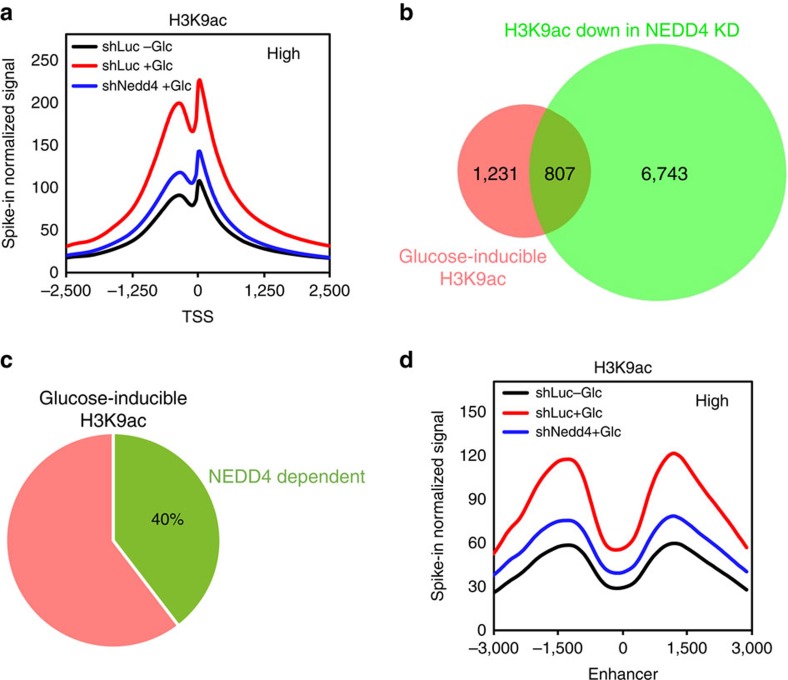
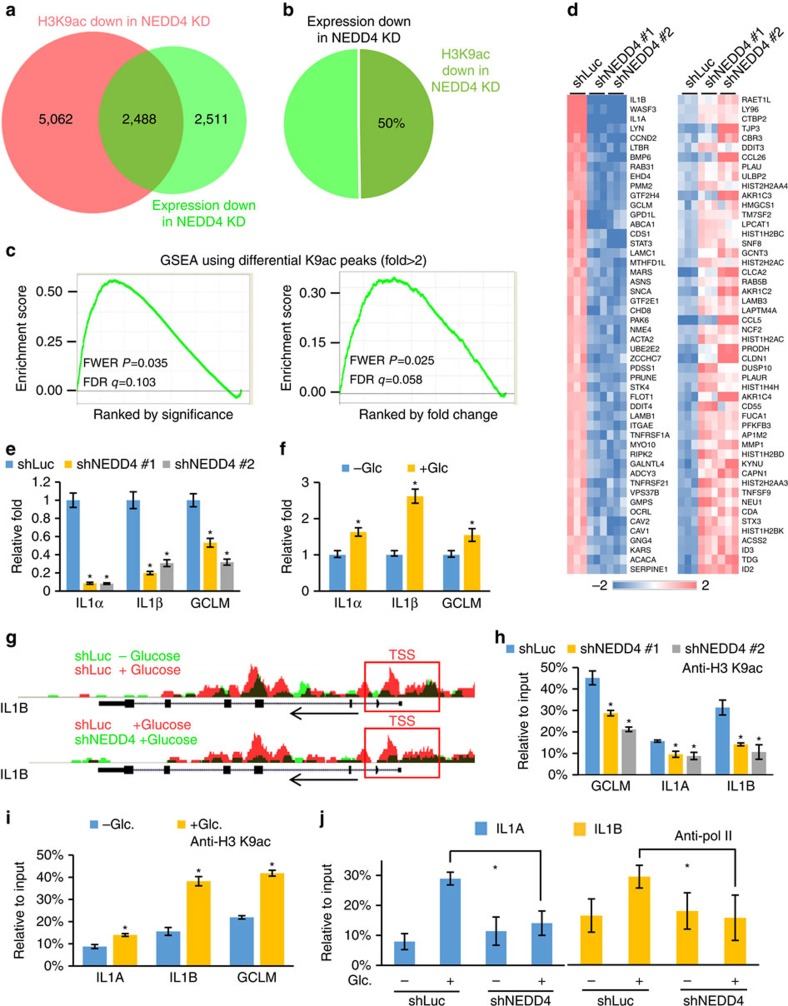
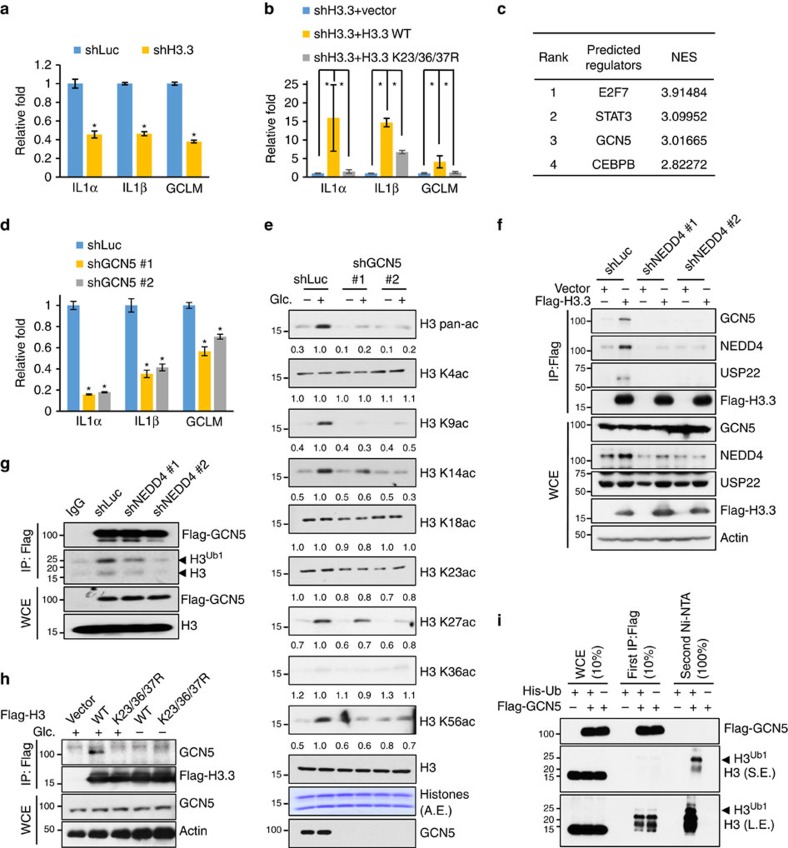
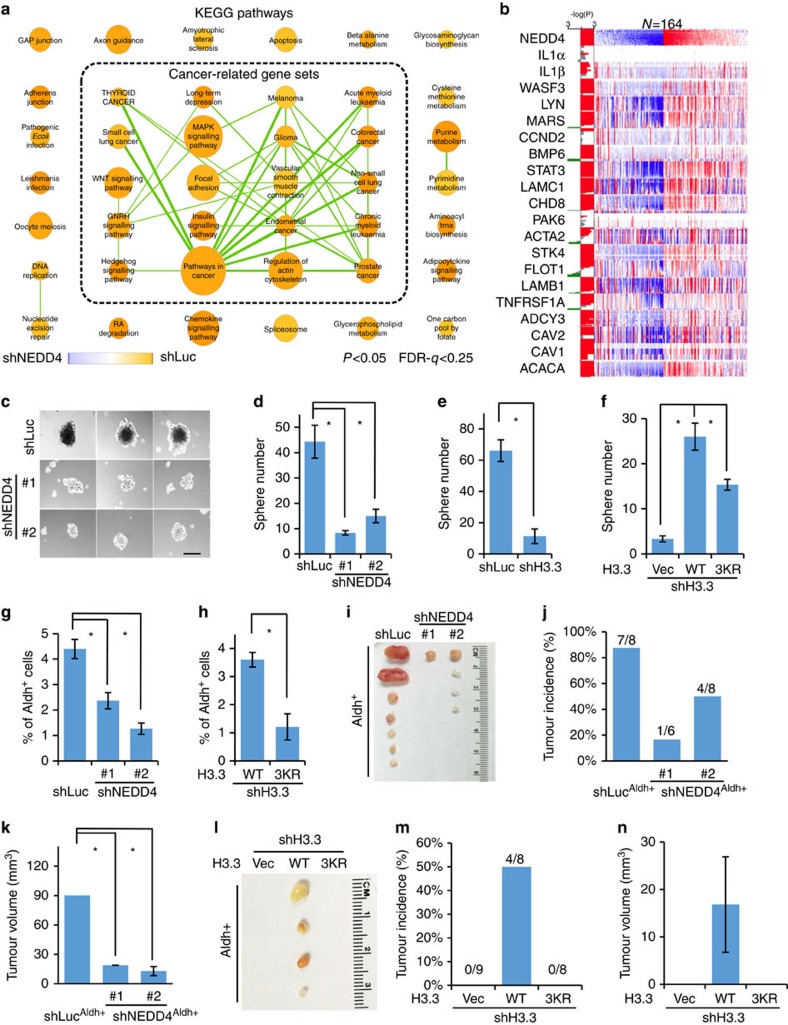
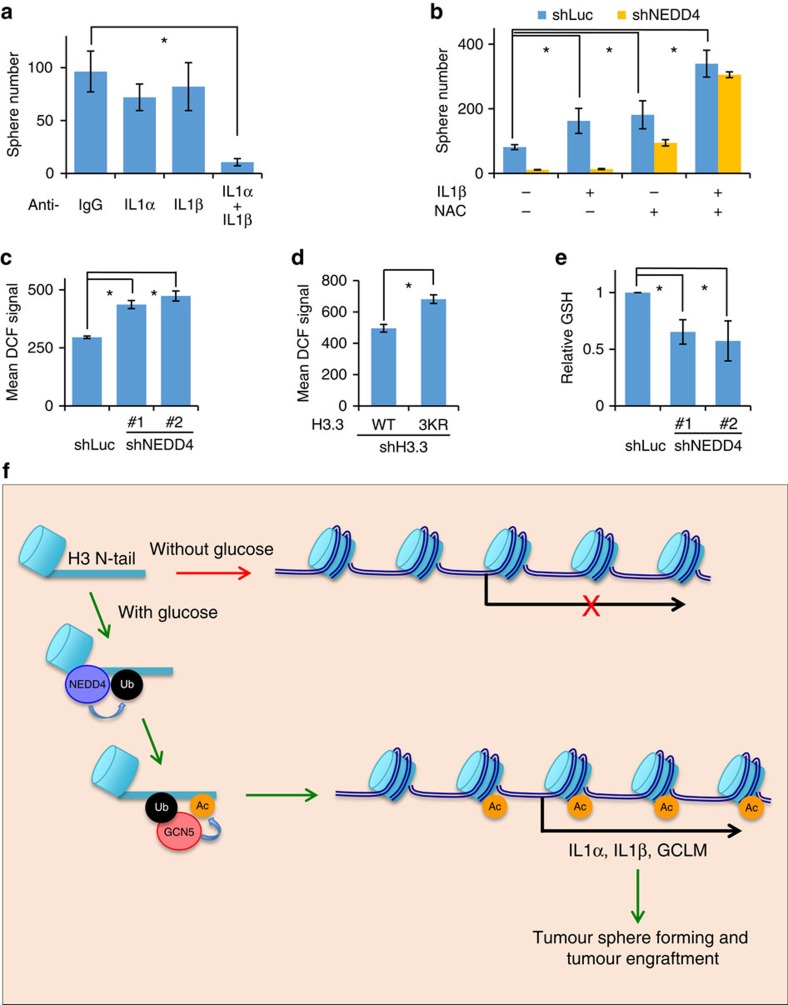
Dynamic changes in histone modifications under various physiological cues play important roles in gene transcription and cancer. Identification of new histone marks critical for cancer development is of particular importance. Here we show that, in a glucose-dependent manner, E3 ubiquitin ligase NEDD4 ubiquitinates histone H3 on lysine 23/36/37 residues, which specifically recruits histone acetyltransferase GCN5 for subsequent H3 acetylation. Genome-wide analysis of chromatin immunoprecipitation followed by sequencing reveals that NEDD4 regulates glucose-induced H3 K9 acetylation at transcription starting site and enhancer regions. Integrative analysis of ChIP-seq and microarray data sets also reveals a consistent role of NEDD4 in transcription activation and H3 K9 acetylation in response to glucose. Functionally, we show that NEDD4-mediated H3 ubiquitination, by transcriptionally activating IL1α, IL1β and GCLM, is important for tumour sphere formation. Together, our study reveals the mechanism for glucose-induced transcriptome reprograming and epigenetic regulation in cancer by inducing NEDD4-dependent H3 ubiquitination.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Regulation of PTEN degradation and NEDD4-1 E3 ligase activity by Numb.Cell Cycle. 2017 May 19;16(10):957-967. doi: 10.1080/15384101.2017.1310351. Epub 2017 Apr 24. Cell Cycle. 2017. PMID: 28437168 Free PMC article.
-
P300-mediated NEDD4 acetylation drives ebolavirus VP40 egress by enhancing NEDD4 ligase activity.PLoS Pathog. 2021 Jun 10;17(6):e1009616. doi: 10.1371/journal.ppat.1009616. eCollection 2021 Jun. PLoS Pathog. 2021. PMID: 34111220 Free PMC article.
-
E3 ubiquitin ligase Nedd4 inhibits AP-1 activity and TNF-α production through targeting p38α for polyubiquitination and subsequent degradation.Sci Rep. 2017 Jul 3;7(1):4521. doi: 10.1038/s41598-017-04072-2. Sci Rep. 2017. PMID: 28674435 Free PMC article.
-
NEDD4 E3 ligase: Functions and mechanism in human cancer.Semin Cancer Biol. 2020 Dec;67(Pt 2):92-101. doi: 10.1016/j.semcancer.2020.03.006. Epub 2020 Apr 11. Semin Cancer Biol. 2020. PMID: 32171886 Review.
-
Epigenetic landscape of amphetamine and methamphetamine addiction in rodents.Epigenetics. 2015;10(7):574-80. doi: 10.1080/15592294.2015.1055441. Epigenetics. 2015. PMID: 26023847 Free PMC article. Review.
Cited by
-
UHRF1 ubiquitin ligase activity supports the maintenance of low-density CpG methylation.Nucleic Acids Res. 2024 Dec 11;52(22):13733-13756. doi: 10.1093/nar/gkae1105. Nucleic Acids Res. 2024. PMID: 39607687 Free PMC article.
-
Key glycometabolism during oocyte maturation and early embryonic development.Reproduction. 2025 Feb 4;169(3):e240275. doi: 10.1530/REP-24-0275. Print 2025 Mar 1. Reproduction. 2025. PMID: 39846956 Free PMC article. Review.
-
Signaling pathways in cancer metabolism: mechanisms and therapeutic targets.Signal Transduct Target Ther. 2023 May 10;8(1):196. doi: 10.1038/s41392-023-01442-3. Signal Transduct Target Ther. 2023. PMID: 37164974 Free PMC article. Review.
-
Chromatin Regulation through Ubiquitin and Ubiquitin-like Histone Modifications.Trends Biochem Sci. 2021 Apr;46(4):258-269. doi: 10.1016/j.tibs.2020.11.005. Epub 2020 Dec 9. Trends Biochem Sci. 2021. PMID: 33308996 Free PMC article. Review.
-
Promising landscape for regulating macrophage polarization: epigenetic viewpoint.Oncotarget. 2017 Apr 11;8(34):57693-57706. doi: 10.18632/oncotarget.17027. eCollection 2017 Aug 22. Oncotarget. 2017. PMID: 28915705 Free PMC article. Review.
References
-
- Graff J. & Tsai L. H. Histone acetylation: molecular mnemonics on the chromatin. Nat. Rev. Neurosci. 14, 97–111 (2013). - PubMed
-
- Brown C. E., Lechner T., Howe L. & Workman J. L. The many HATs of transcription coactivators. Trends Biochem. Sci. 25, 15–19 (2000). - PubMed
-
- Lee K. K. & Workman J. L. Histone acetyltransferase complexes: one size doesn't fit all. Nat. Rev. Mol. Cell Biol. 8, 284–295 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
