

Exonuclease processivity of archaeal replicative DNA polymerase in association with PCNA is expedited by mismatches in DNA
- PMID: 28300173
- PMCID: PMC5353730
- DOI: 10.1038/srep44582
Exonuclease processivity of archaeal replicative DNA polymerase in association with PCNA is expedited by mismatches in DNA
Abstract
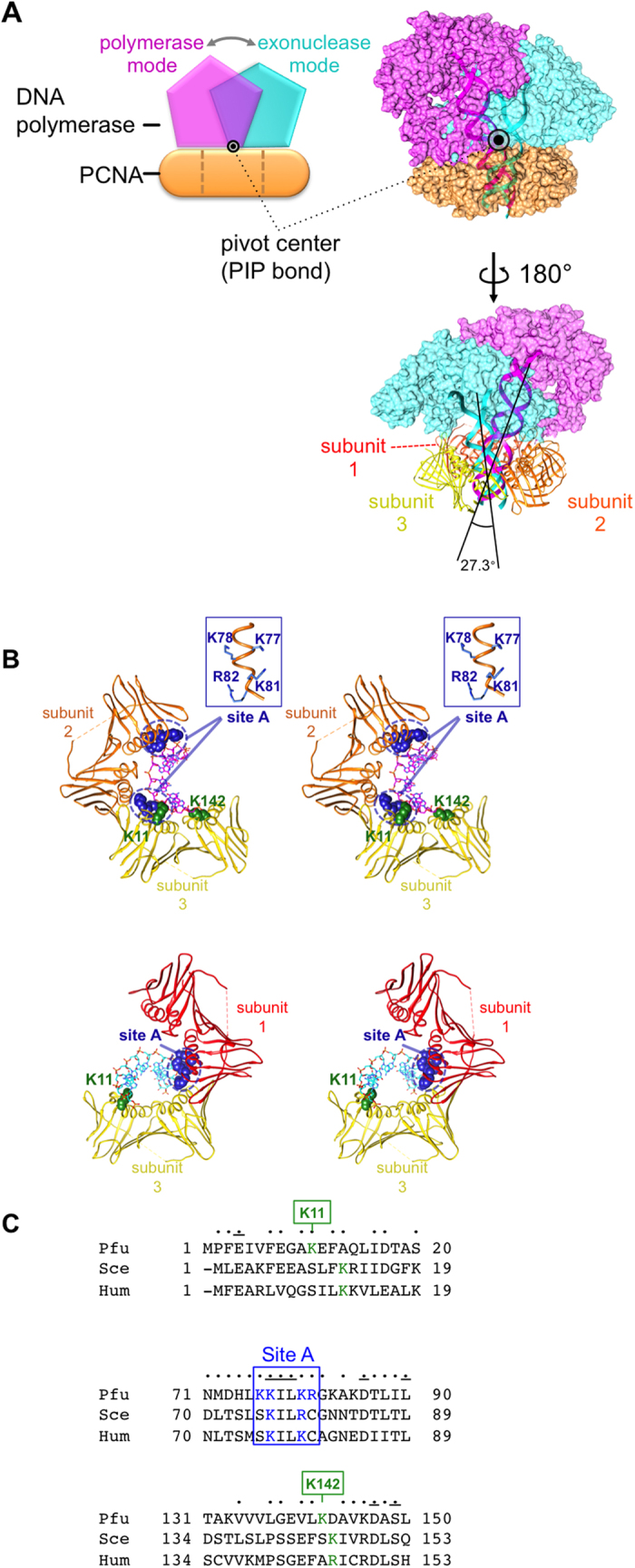
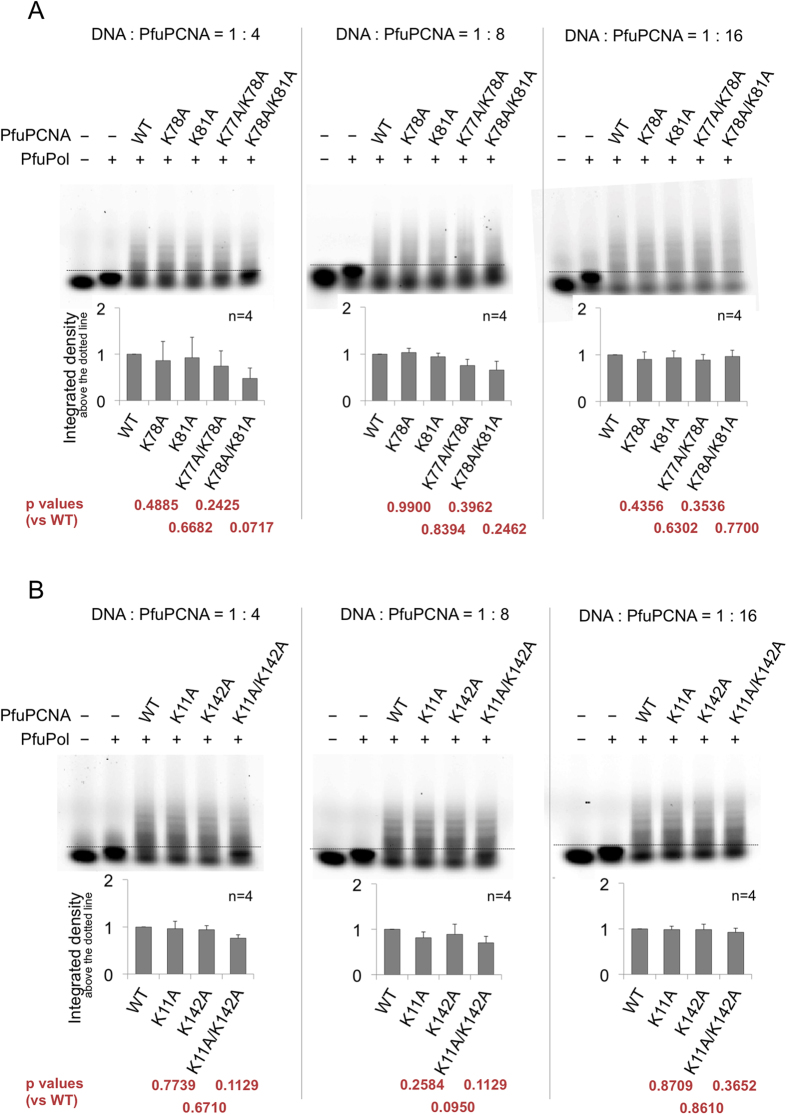
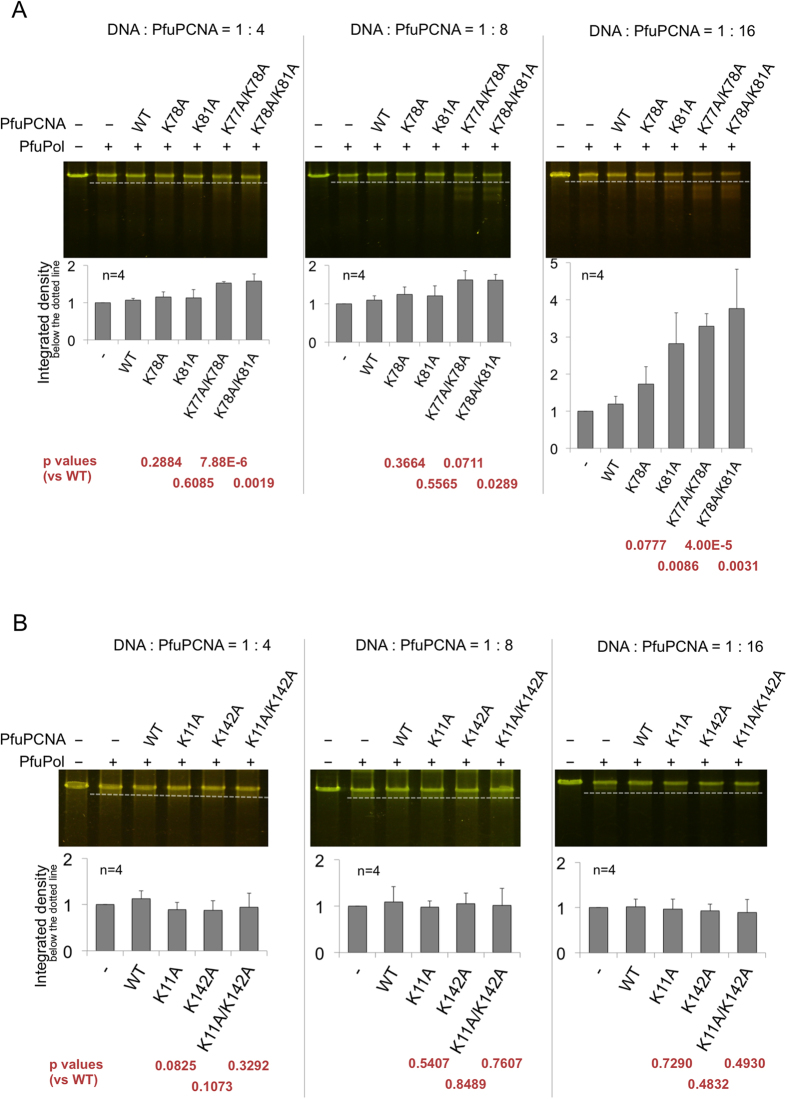
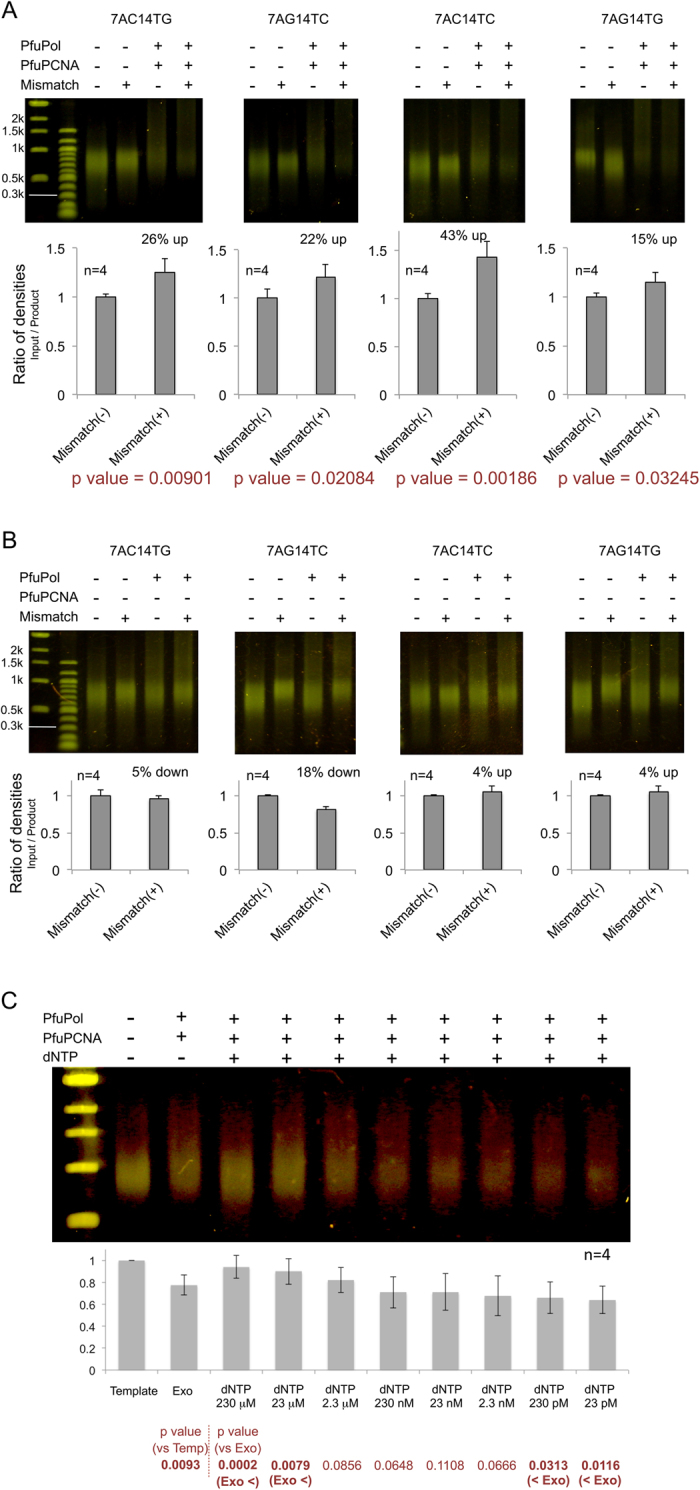
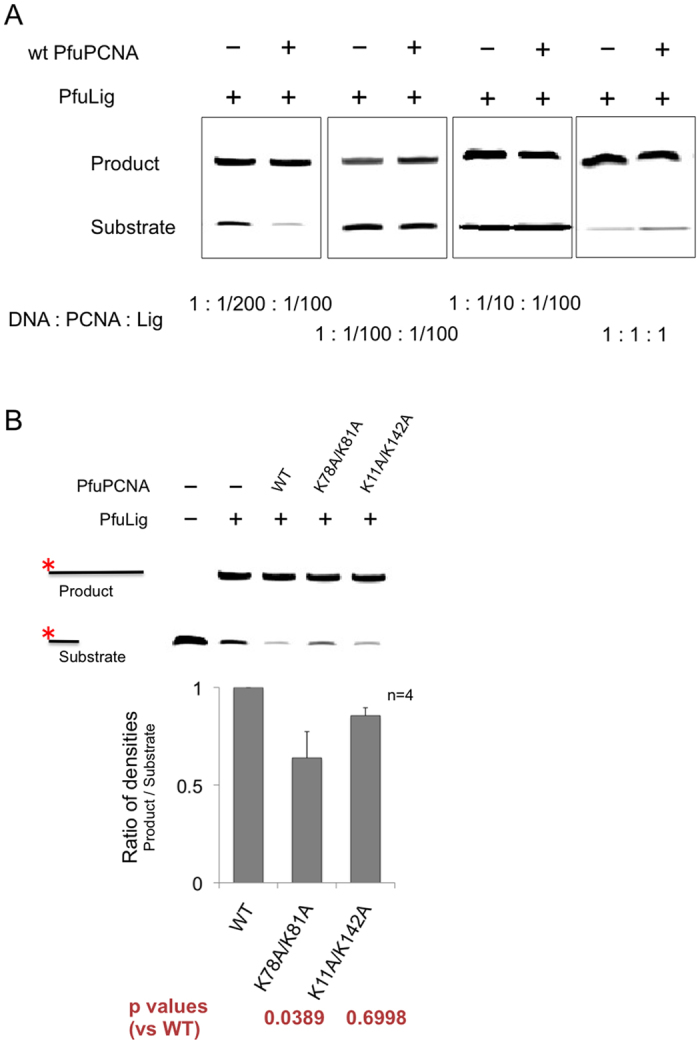
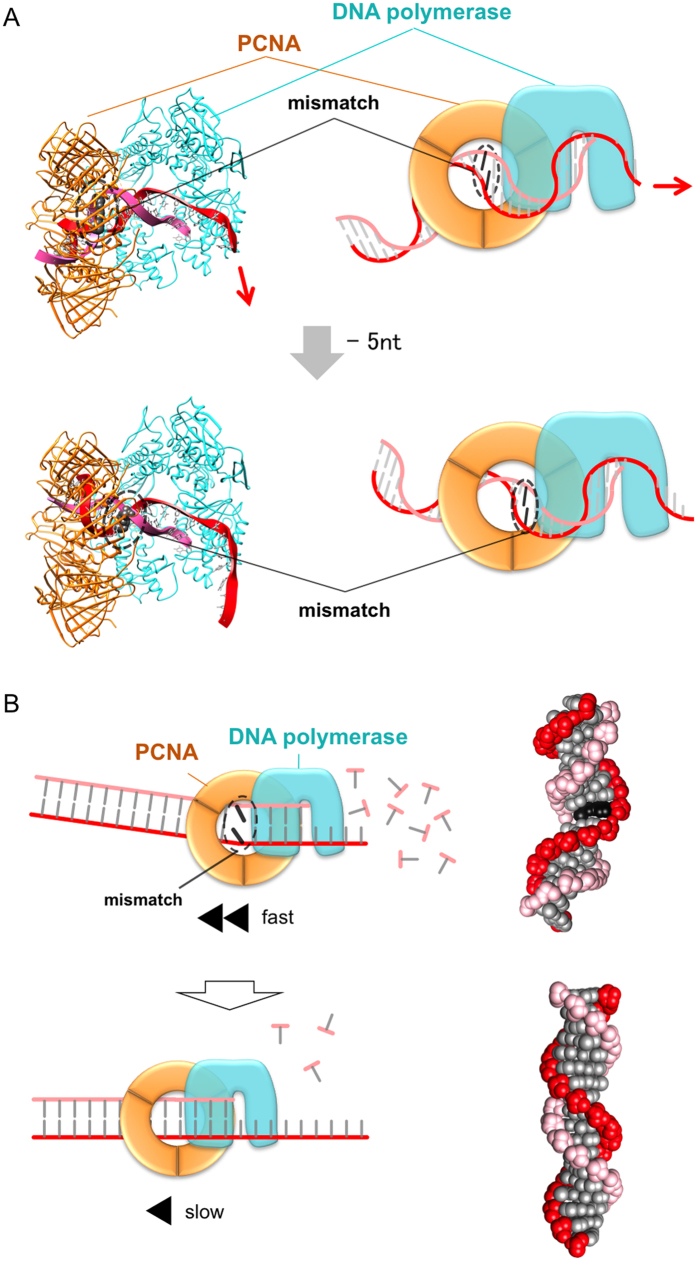
Family B DNA polymerases comprise polymerase and 3' ->5' exonuclease domains, and detect a mismatch in a newly synthesized strand to remove it in cooperation with Proliferating cell nuclear antigen (PCNA), which encircles the DNA to provide a molecular platform for efficient protein-protein and protein-DNA interactions during DNA replication and repair. Once the repair is completed, the enzyme must stop the exonucleolytic process and switch to the polymerase mode. However, the cue to stop the degradation is unclear. We constructed several PCNA mutants and found that the exonuclease reaction was enhanced in the mutants lacking the conserved basic patch, located on the inside surface of PCNA. These mutants may mimic the Pol/PCNA complex processing the mismatched DNA, in which PCNA cannot interact rigidly with the irregularly distributed phosphate groups outside the dsDNA. Indeed, the exonuclease reaction with the wild type PCNA was facilitated by mismatched DNA substrates. PCNA may suppress the exonuclease reaction after the removal of the mismatched nucleotide. PCNA seems to act as a "brake" that stops the exonuclease mode of the DNA polymerase after the removal of a mismatched nucleotide from the substrate DNA, for the prompt switch to the DNA polymerase mode.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Tsurimoto T. PCNA, a multifunctional ring on DNA. Biochim. Biophys. Acta 1443, 23–39 (1998). - PubMed
-
- Kelman Z. & Hurwitz J. Protein-PCNA interactions: a DNA scanning mechanism? Trends Biochem. Sci. 23, 236–238 (1998). - PubMed
-
- Maga G. & Hübscher U. Proliferating cell nuclear antigen (PCNA): a dancer with many partners. J. Cell Sci. 116, 3051–3060 (2003). - PubMed
-
- Krishna T. S. R., Kong X.-P., Gary S., Burgers P. M. & Kuriyan J. Crystal structure of the eukaryotic DNA polymerase processivity factor PCNA. Cell 79, 1233–1243 (1994). - PubMed
-
- Gulbis J. M., Kelman Z., Hurwitz J., O’Donnell M. & Kuriyan J. Structure of the C-terminal region of p21WAF1/CIP1 complexed with human PCNA. Cell 87, 297–306 (1996). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
