

Regulation of RhoA activity by the cellular prion protein
- PMID: 28300846
- PMCID: PMC5386549
- DOI: 10.1038/cddis.2017.37
Regulation of RhoA activity by the cellular prion protein
Abstract
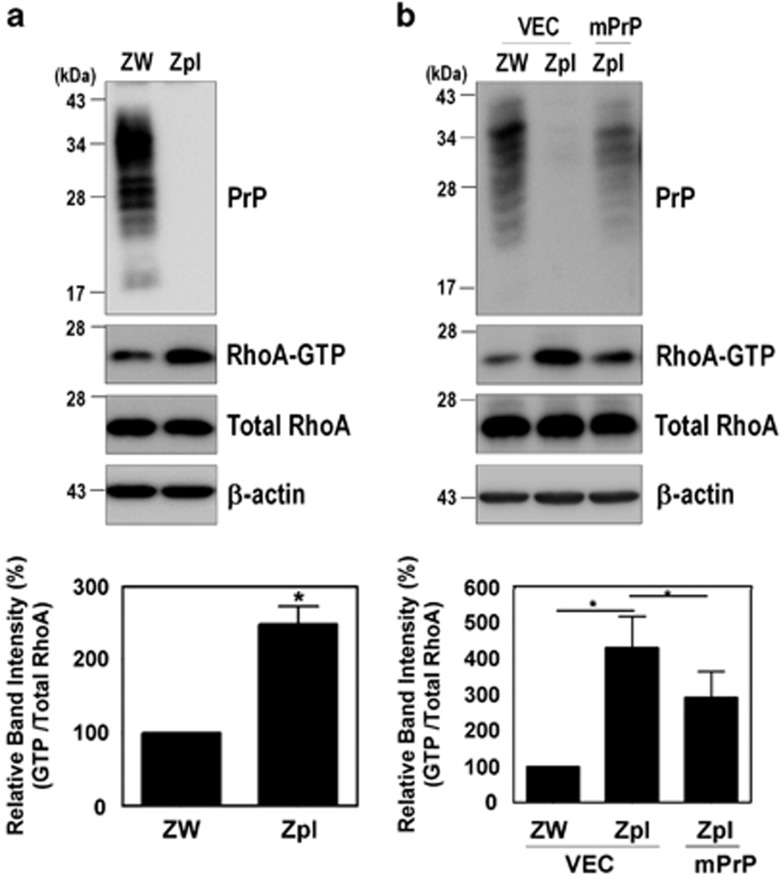
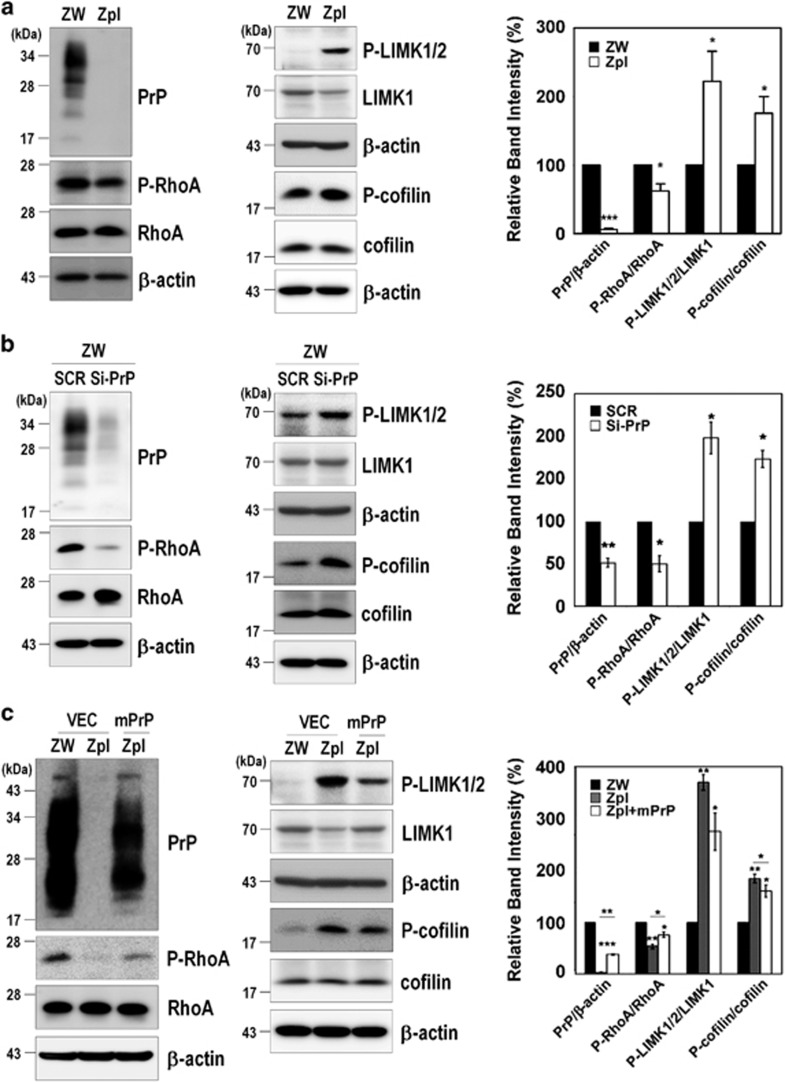
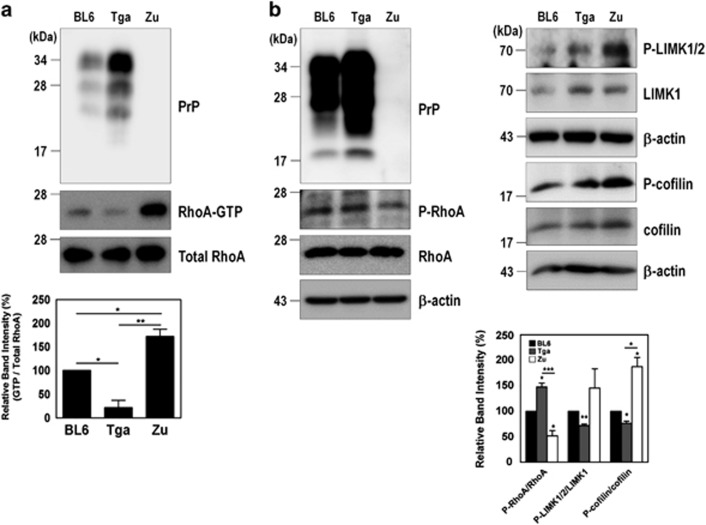
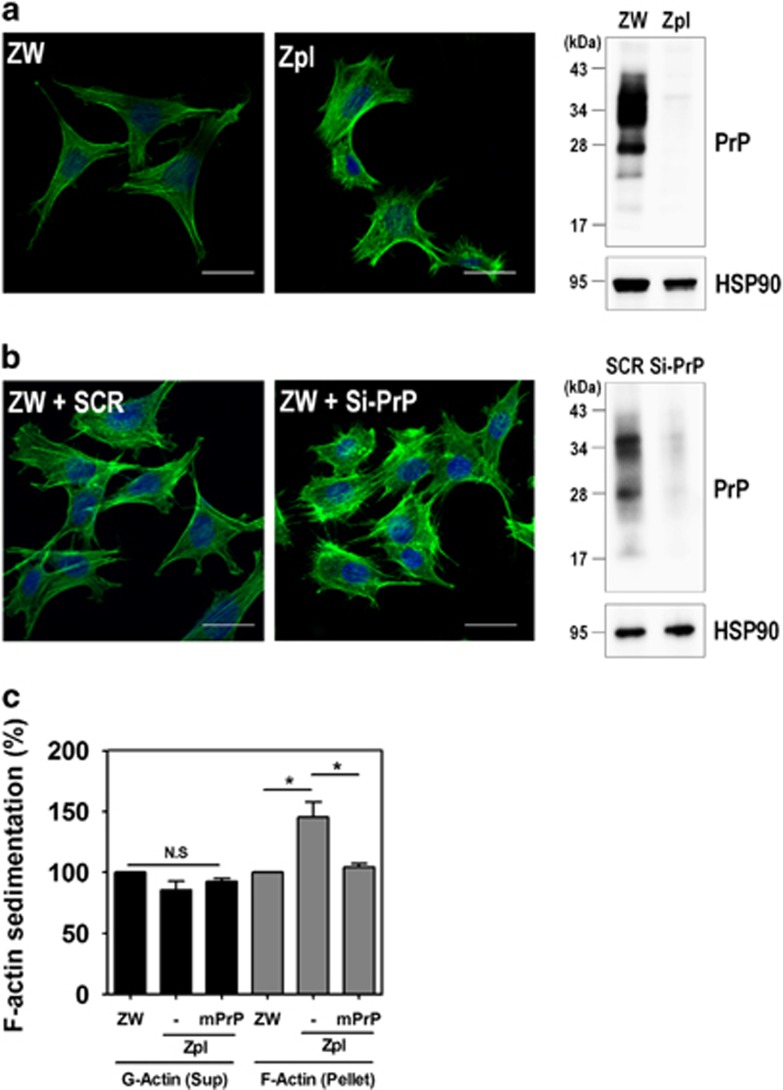
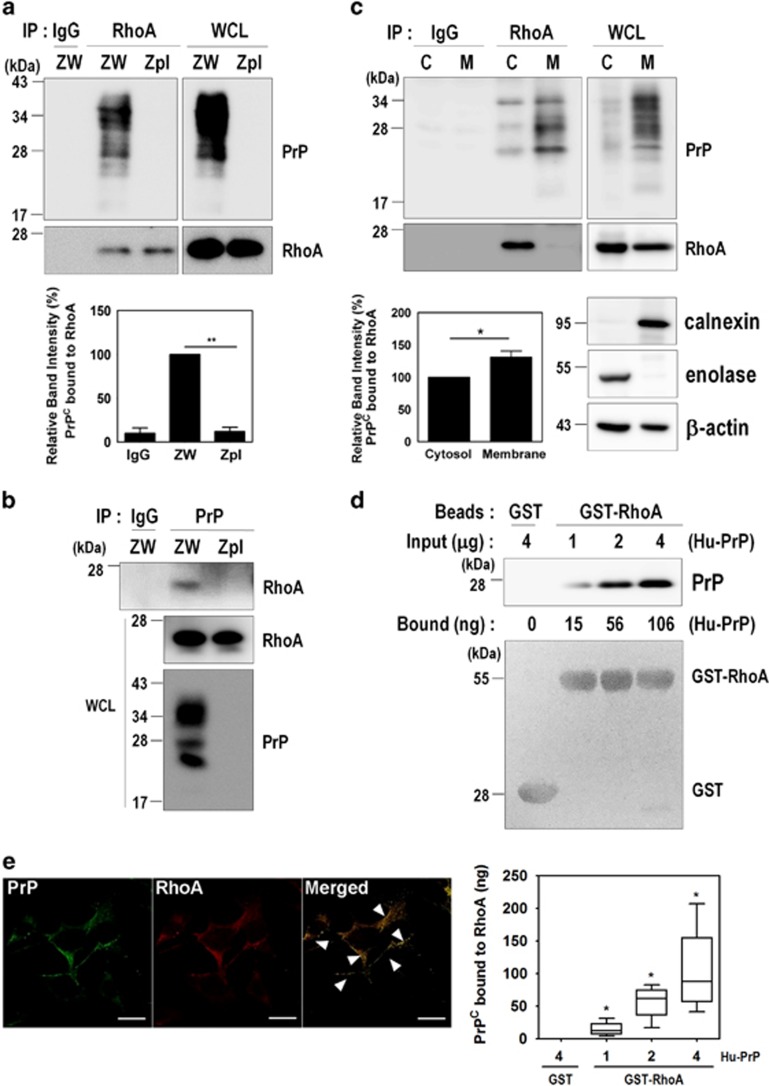
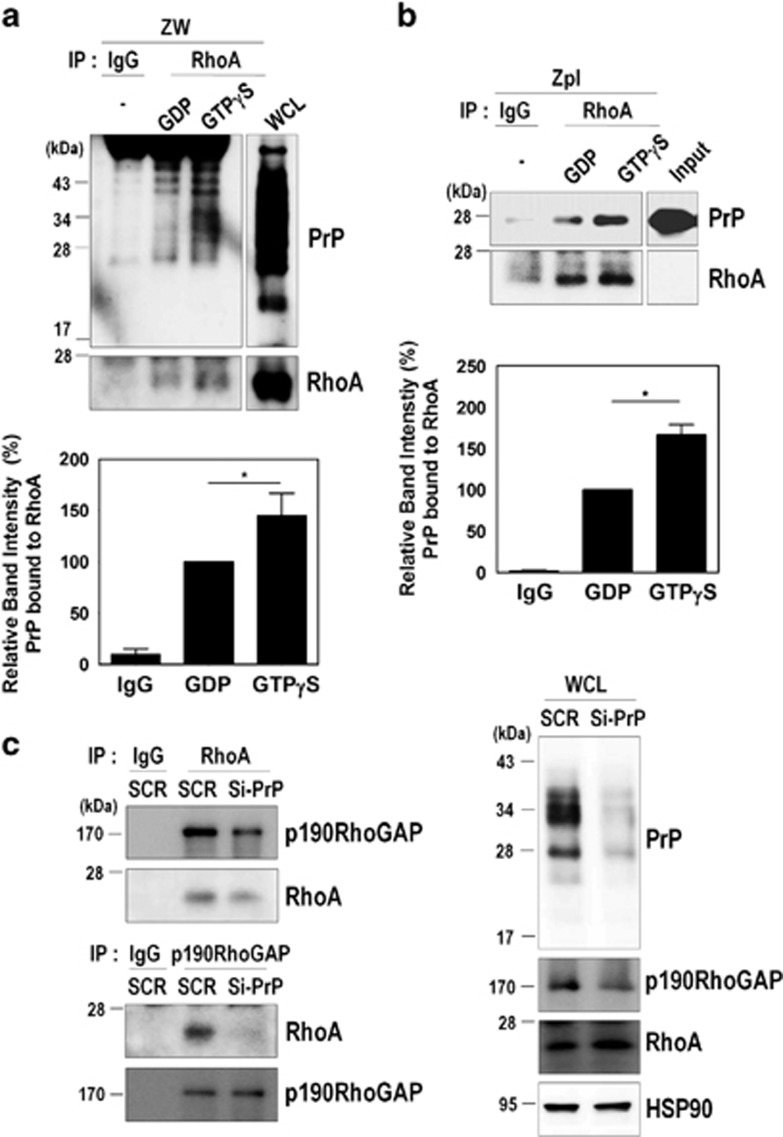
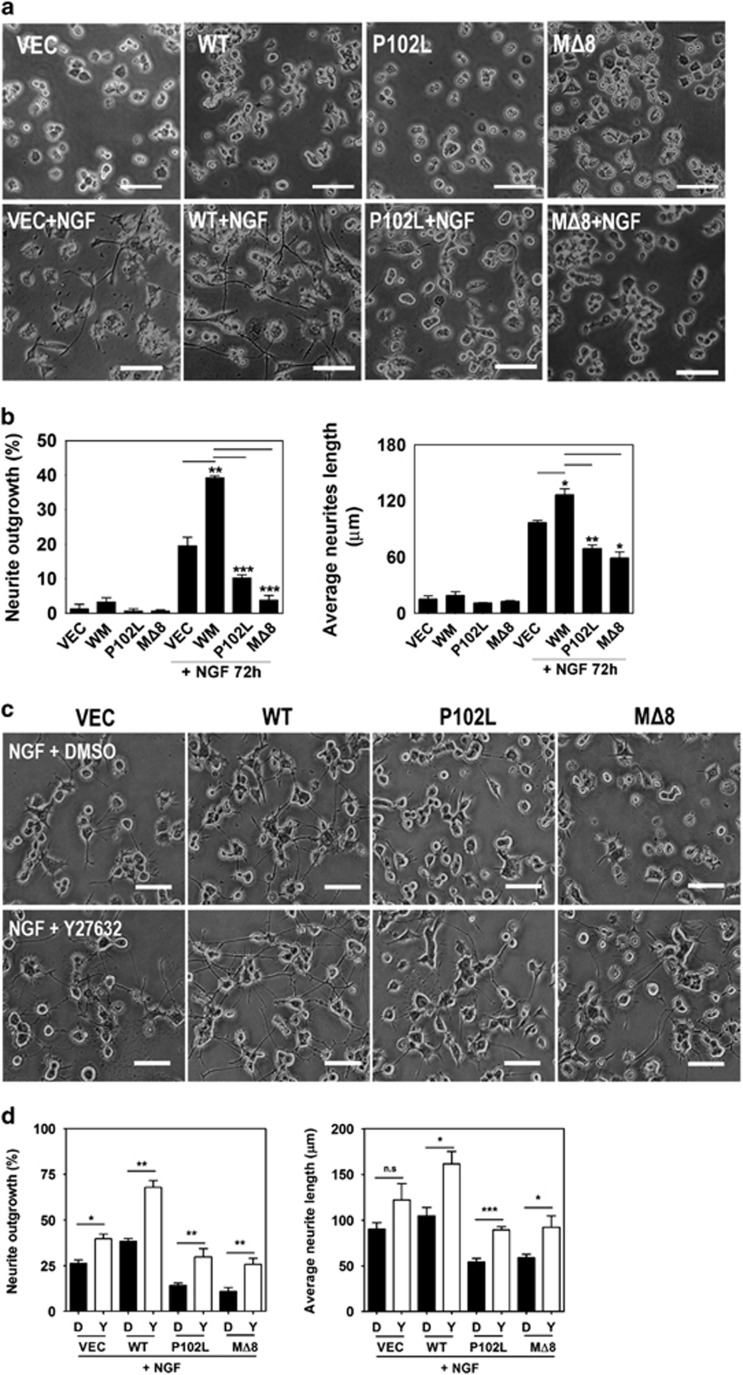
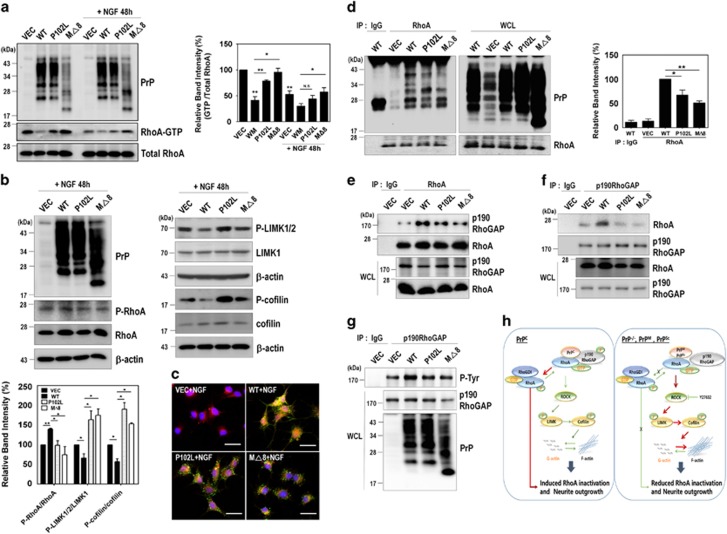
The cellular prion protein (PrPC) is a highly conserved glycosylphosphatidylinositol (GPI)-anchored membrane protein that is involved in the signal transduction during the initial phase of neurite outgrowth. The Ras homolog gene family member A (RhoA) is a small GTPase that is known to have an essential role in regulating the development, differentiation, survival, and death of neurons in the central nervous system. Although recent studies have shown the dysregulation of RhoA in a variety of neurodegenerative diseases, the role of RhoA in prion pathogenesis remains unclear. Here, we investigated the regulation of RhoA-mediated signaling by PrPC using both in vitro and in vivo models and found that overexpression of PrPC significantly induced RhoA inactivation and RhoA phosphorylation in hippocampal neuronal cells and in the brains of transgenic mice. Using siRNA-mediated depletion of endogenous PrPC and overexpression of disease-associated mutants of PrPC, we confirmed that PrPC induced RhoA inactivation, which accompanied RhoA phosphorylation but reduced the phosphorylation levels of LIM kinase (LIMK), leading to cofilin activation. In addition, PrPC colocalized with RhoA, and the overexpression of PrPC significantly increased neurite outgrowth in nerve growth factor-treated PC12 cells through RhoA inactivation. However, the disease-associated mutants of PrPC decreased neurite outgrowth compared with wild-type PrPC. Moreover, inhibition of Rho-associated kinase (ROCK) substantially facilitated neurite outgrowth in NGF-treated PC12 cells, similar to the effect induced by PrPC. Interestingly, we found that the induction of RhoA inactivation occurred through the interaction of PrPC with RhoA and that PrPC enhanced the interaction between RhoA and p190RhoGAP (a GTPase-activating protein). These findings suggest that the interactions of PrPC with RhoA and p190RhoGAP contribute to neurite outgrowth by controlling RhoA inactivation and RhoA-mediated signaling and that disease-associated mutations of PrPC impair RhoA inactivation, which in turn leads to prion-related neurodegeneration.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Burridge K, Wennerberg K. Rho and Rac take center stage. Cell 2004; 116: 167–179. - PubMed
-
- Kim HJ, Kim JG, Moon MY, Park SH, Park JB. IκB kinase gamma/nuclear factor-κB-essential modulator (IKKgamma/NEMO) facilitates RhoA GTPase activation, which, in turn, activates Rho-associated KINASE (ROCK) to phosphorylate IKKbeta in response to transforming growth factor TGF-β1. J Biol Chem 2014; 289: 1429–1440. - PMC - PubMed
-
- Kjoller L, Hall A. Signaling to Rho GTPases. Exp Cell Res 1999; 253: 166–179. - PubMed
-
- Govek EE, Newey SE, Van Aelst L. The role of the Rho GTPases in neuronal development. Genes Dev 2005; 19: 1–49. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
