

Dissection of TALE-dependent gene activation reveals that they induce transcription cooperatively and in both orientations
- PMID: 28301511
- PMCID: PMC5354296
- DOI: 10.1371/journal.pone.0173580
Dissection of TALE-dependent gene activation reveals that they induce transcription cooperatively and in both orientations
Erratum in
-
Correction: Dissection of TALE-dependent gene activation reveals that they induce transcription cooperatively and in both orientations.PLoS One. 2017 Apr 6;12(4):e0175653. doi: 10.1371/journal.pone.0175653. eCollection 2017. PLoS One. 2017. PMID: 28384283 Free PMC article.
Abstract
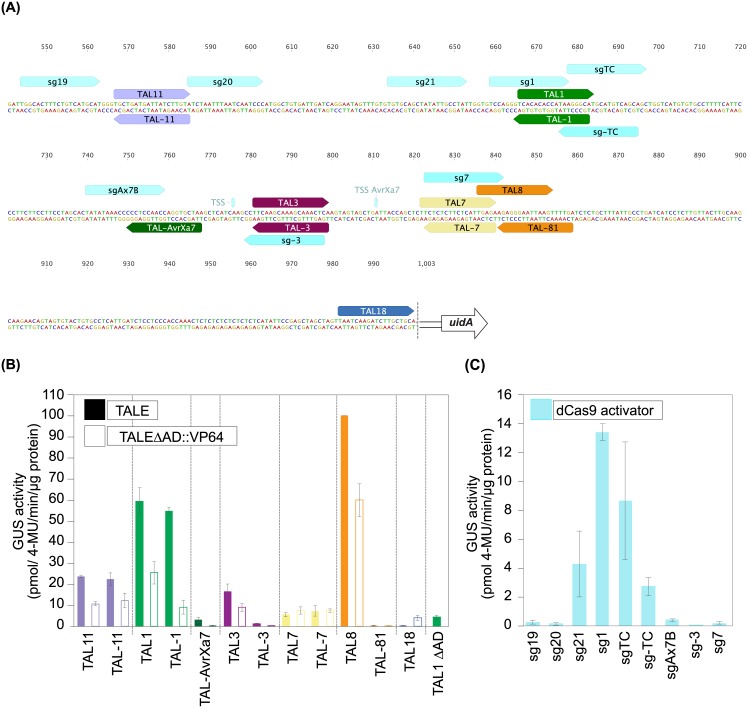
Plant-pathogenic Xanthomonas bacteria inject transcription activator-like effector proteins (TALEs) into host cells to specifically induce transcription of plant genes and enhance susceptibility. Although the DNA-binding mode is well-understood it is still ambiguous how TALEs initiate transcription and whether additional promoter elements are needed to support this. To systematically dissect prerequisites for transcriptional initiation the activity of one TALE was compared on different synthetic Bs4 promoter fragments. In addition, a large collection of artificial TALEs spanning the OsSWEET14 promoter was compared. We show that the presence of a TALE alone is not sufficient to initiate transcription suggesting the requirement of additional supporting promoter elements. At the OsSWEET14 promoter TALEs can initiate transcription from various positions, in a synergistic manner of multiple TALEs binding in parallel to the promoter, and even by binding in reverse orientation. TALEs are known to shift the transcriptional start site, but our data show that this shift depends on the individual position of a TALE within a promoter context. Our results implicate that TALEs function like classical enhancer-binding proteins and initiate transcription in both orientations which has consequences for in planta target gene prediction and design of artificial activators.
Conflict of interest statement
Figures






References
-
- Boch J, Bonas U, Lahaye T. TAL effectors—pathogen strategies and plant resistance engineering. New Phytol. 2014;204: 823–832. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
