

Generation of murine tumor cell lines deficient in MHC molecule surface expression using the CRISPR/Cas9 system
- PMID: 28301575
- PMCID: PMC5354463
- DOI: 10.1371/journal.pone.0174077
Generation of murine tumor cell lines deficient in MHC molecule surface expression using the CRISPR/Cas9 system
Erratum in
-
Correction: Generation of murine tumor cell lines deficient in MHC molecule surface expression using the CRISPR/Cas9 system.PLoS One. 2018 Dec 19;13(12):e0209719. doi: 10.1371/journal.pone.0209719. eCollection 2018. PLoS One. 2018. PMID: 30566524 Free PMC article.
Abstract
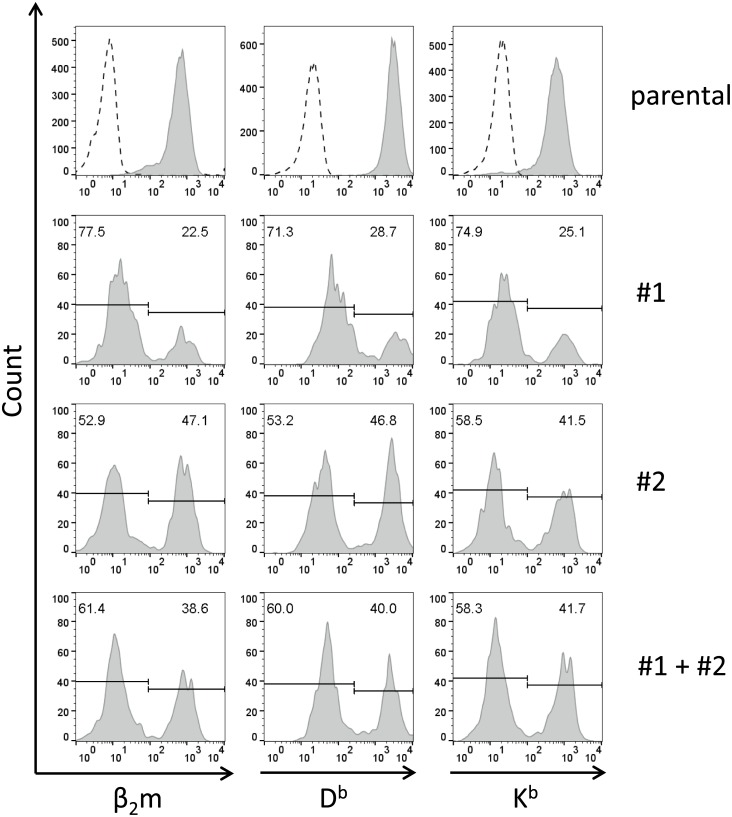
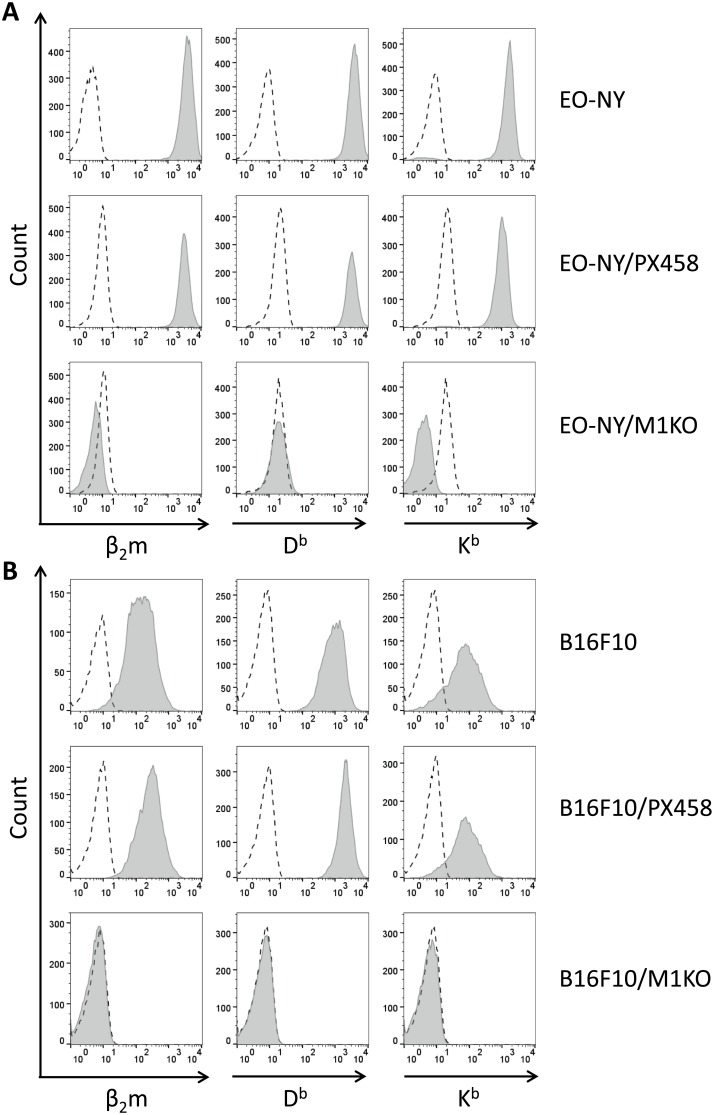
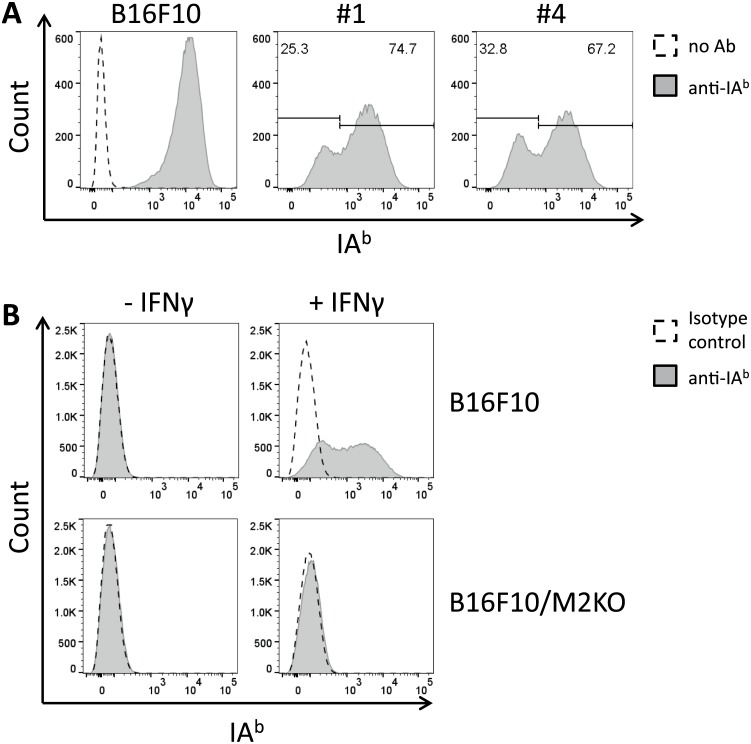
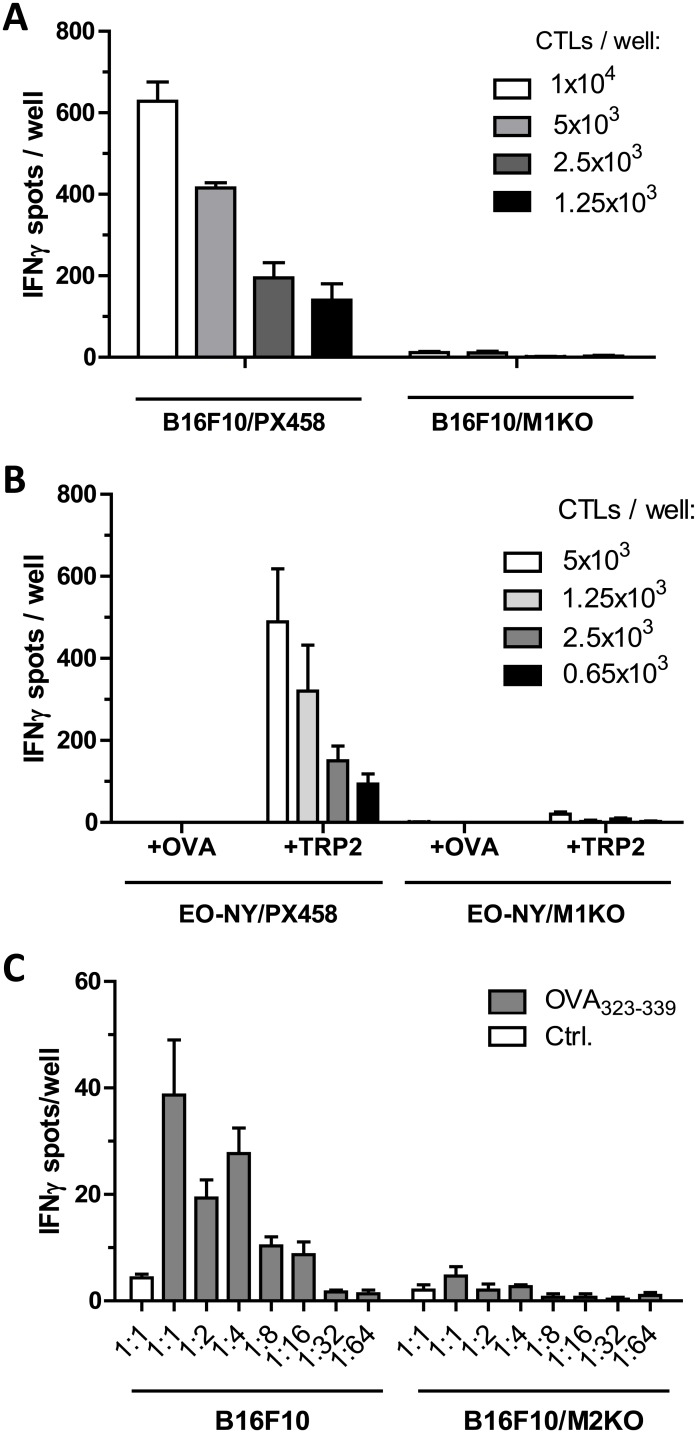
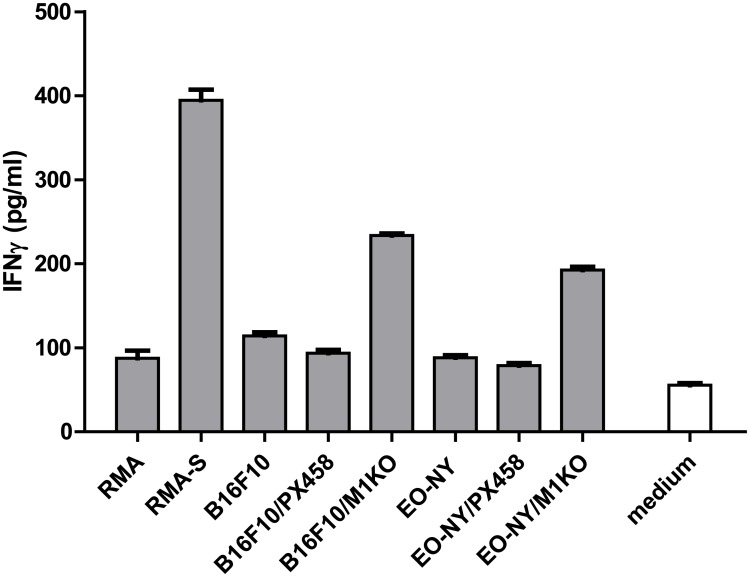
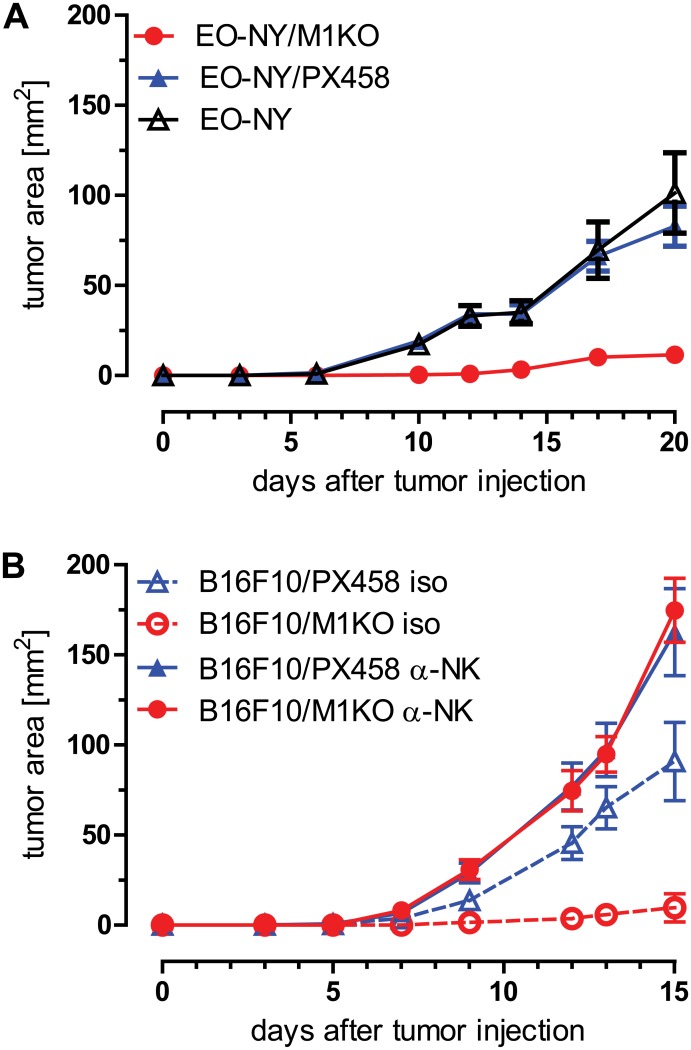
In this study, the CRISPR/Cas9 technology was used to establish murine tumor cell lines, devoid of MHC I or MHC II surface expression, respectively. The melanoma cell line B16F10 and the murine breast cancer cell line EO-771, the latter stably expressing the tumor antigen NY-BR-1 (EO-NY), were transfected with an expression plasmid encoding a β2m-specific single guide (sg)RNA and Cas9. The resulting MHC I negative cells were sorted by flow cytometry to obtain single cell clones, and loss of susceptibility of peptide pulsed MHC I negative clones to peptide-specific CTL recognition was determined by IFNγ ELISpot assay. The β2m knockout (KO) clones did not give rise to tumors in syngeneic mice (C57BL/6N), unless NK cells were depleted, suggesting that outgrowth of the β2m KO cell lines was controlled by NK cells. Using sgRNAs targeting the β-chain encoding locus of the IAb molecule we also generated several B16F10 MHC II KO clones. Peptide loaded B16F10 MHC II KO cells were insusceptible to recognition by OT-II cells and tumor growth was unaltered compared to parental B16F10 cells. Thus, in our hands the CRISPR/Cas9 system has proven to be an efficient straight forward strategy for the generation of MHC knockout cell lines. Such cell lines could serve as parental cells for co-transfection of compatible HLA alleles together with human tumor antigens of interest, thereby facilitating the generation of HLA matched transplantable tumor models, e.g. in HLAtg mouse strains of the newer generation, lacking cell surface expression of endogenous H2 molecules. In addition, our tumor cell lines established might offer a useful tool to investigate tumor reactive T cell responses that function independently from MHC molecule surface expression by the tumor.
Conflict of interest statement
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
