

Reconstructing Ancient Proteins to Understand the Causes of Structure and Function
- PMID: 28301769
- PMCID: PMC6141191
- DOI: 10.1146/annurev-biophys-070816-033631
Reconstructing Ancient Proteins to Understand the Causes of Structure and Function
Abstract
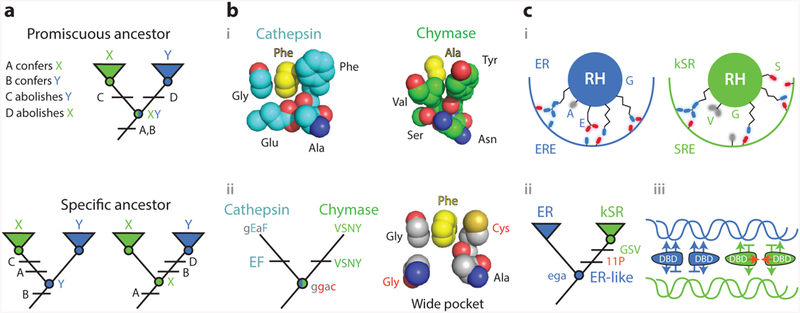
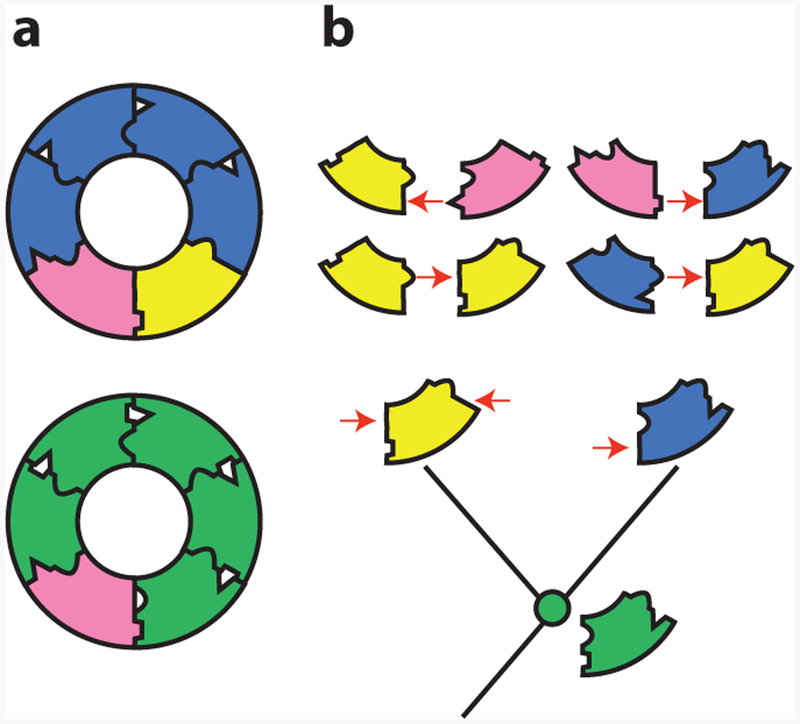
A central goal in biochemistry is to explain the causes of protein sequence, structure, and function. Mainstream approaches seek to rationalize sequence and structure in terms of their effects on function and to identify function's underlying determinants by comparing related proteins to each other. Although productive, both strategies suffer from intrinsic limitations that have left important aspects of many proteins unexplained. These limits can be overcome by reconstructing ancient proteins, experimentally characterizing their properties, and retracing their evolution through time. This approach has proven to be a powerful means for discovering how historical changes in sequence produced the functions, structures, and other physical/chemical characteristics of modern proteins. It has also illuminated whether protein features evolved because of functional optimization, historical constraint, or blind chance. Here we review recent studies employing ancestral protein reconstruction and show how they have produced new knowledge not only of molecular evolutionary processes but also of the underlying determinants of modern proteins' physical, chemical, and biological properties.
Keywords: ancestral reconstruction; epistasis; evolutionary biochemistry; historical contingency; vertical analysis.
Figures
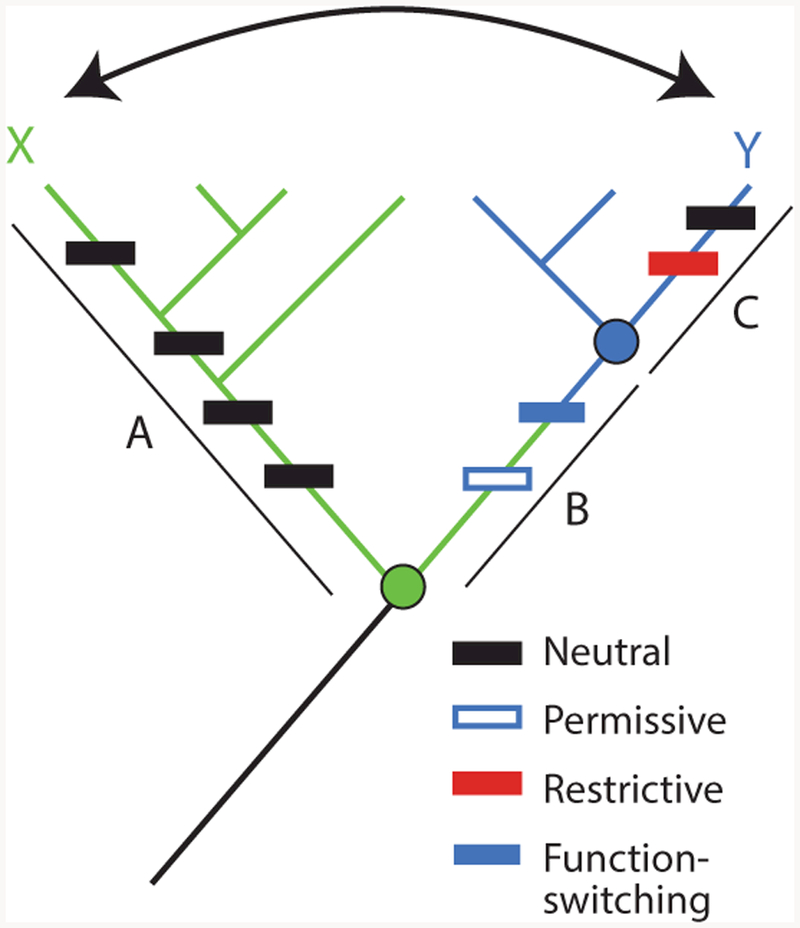
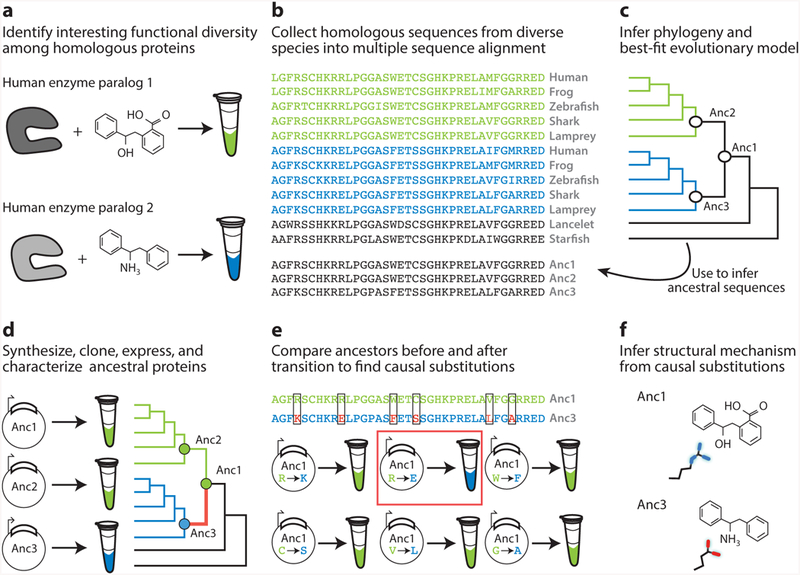
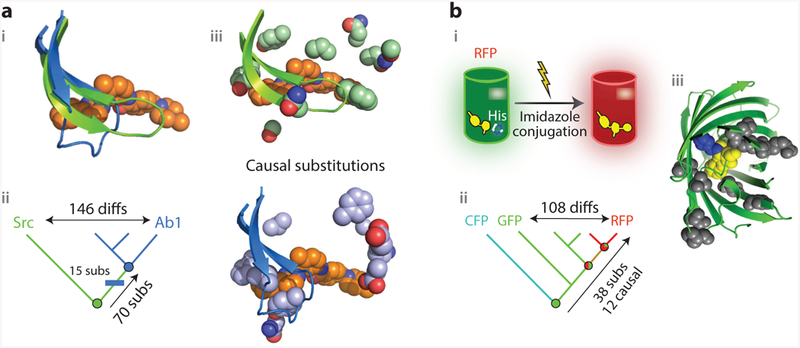
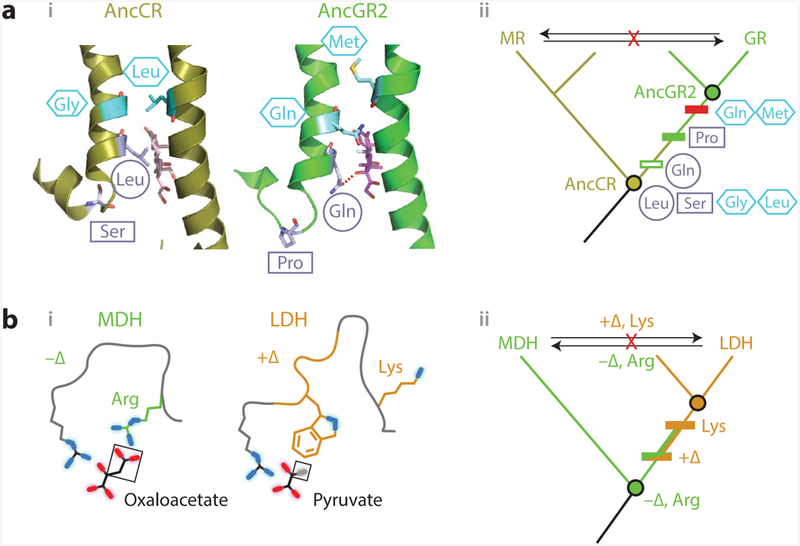
References
-
- Abascal F, Zardoya R, Posada D. 2005. ProtTest: selection of best-fit models of protein evolution. Bioinformatics 21:2104–5 - PubMed
-
- Aharoni A, Gaidukov L, Khersonsky O, McQ Gould S, Roodveldt C, Tawfik DS. 2005. The ‘evolvability’ of promiscuous protein functions. Nat. Genet 37:73–76 - PubMed
-
- Ahnert SE, Marsh JA, Hernandez H, Robinson CV, Teichmann SA. 2015. Principles of assembly reveal a periodic table of protein complexes. Science 350:aaa2245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
