

Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition
- PMID: 28302675
- PMCID: PMC5435430
- DOI: 10.1105/tpc.16.00891
Receptor Kinases in Plant-Pathogen Interactions: More Than Pattern Recognition
Abstract
Receptor-like kinases (RLKs) and Receptor-like proteins (RLPs) play crucial roles in plant immunity, growth, and development. Plants deploy a large number of RLKs and RLPs as pattern recognition receptors (PRRs) that detect microbe- and host-derived molecular patterns as the first layer of inducible defense. Recent advances have uncovered novel PRRs, their corresponding ligands, and mechanisms underlying PRR activation and signaling. In general, PRRs associate with other RLKs and function as part of multiprotein immune complexes at the cell surface. Innovative strategies have emerged for the rapid identification of microbial patterns and their cognate PRRs. Successful pathogens can evade or block host recognition by secreting effector proteins to "hide" microbial patterns or inhibit PRR-mediated signaling. Furthermore, newly identified pathogen effectors have been shown to manipulate RLKs controlling growth and development by mimicking peptide hormones of host plants. The ongoing studies illustrate the importance of diverse plant RLKs in plant disease resistance and microbial pathogenesis.
© 2017 American Society of Plant Biologists. All rights reserved.
Figures
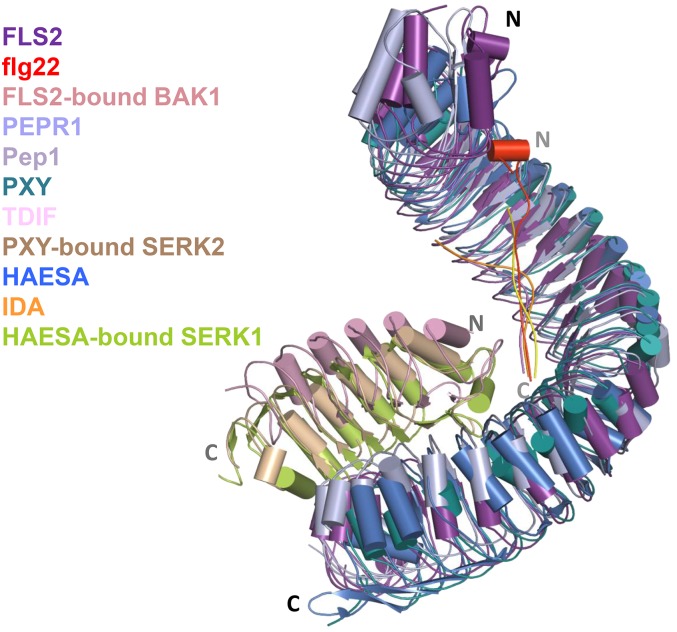
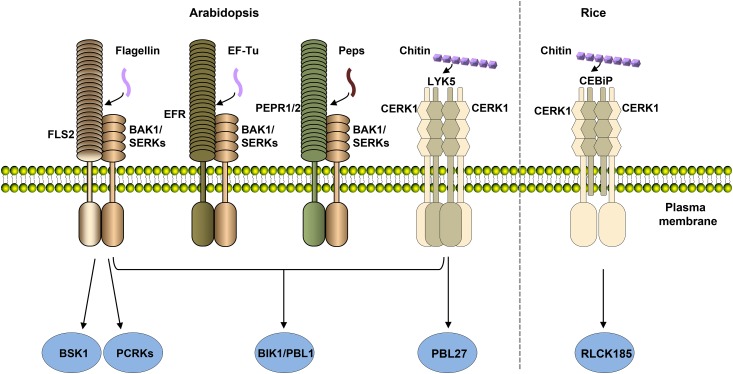
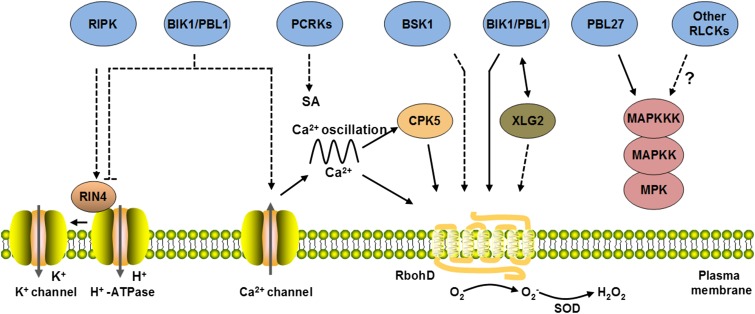
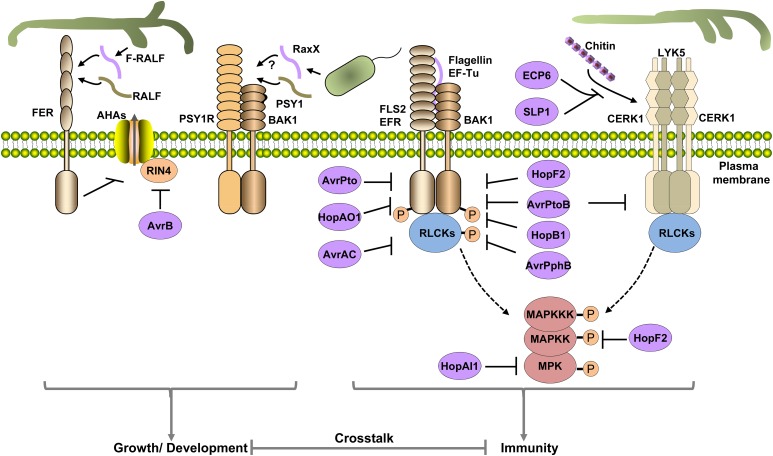
References
-
- Albert I., et al. (2015). An RLP23-SOBIR1-BAK1 complex mediates NLP-triggered immunity. Nat. Plants 1: 15140. - PubMed
-
- Alcázar R., García A.V., Kronholm I., de Meaux J., Koornneef M., Parker J.E., Reymond M. (2010). Natural variation at Strubbelig Receptor Kinase 3 drives immune-triggered incompatibilities between Arabidopsis thaliana accessions. Nat. Genet. 42: 1135–1139. - PubMed
-
- Ao Y., Li Z., Feng D., Xiong F., Liu J., Li J.F., Wang M., Wang J., Liu B., Wang H.B. (2014). OsCERK1 and OsRLCK176 play important roles in peptidoglycan and chitin signaling in rice innate immunity. Plant J. 80: 1072–1084. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
