

Cardiac Regeneration: Lessons From Development
- PMID: 28302741
- PMCID: PMC5358810
- DOI: 10.1161/CIRCRESAHA.116.309040
Cardiac Regeneration: Lessons From Development
Abstract
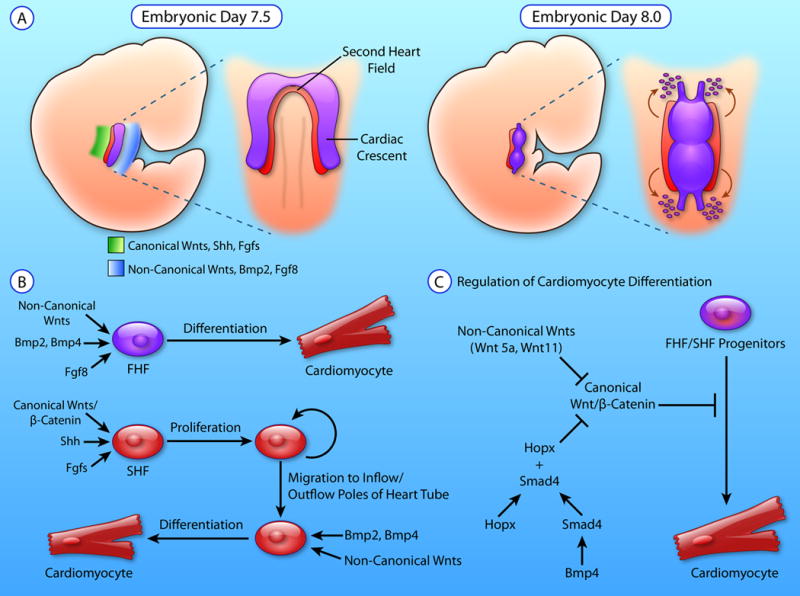
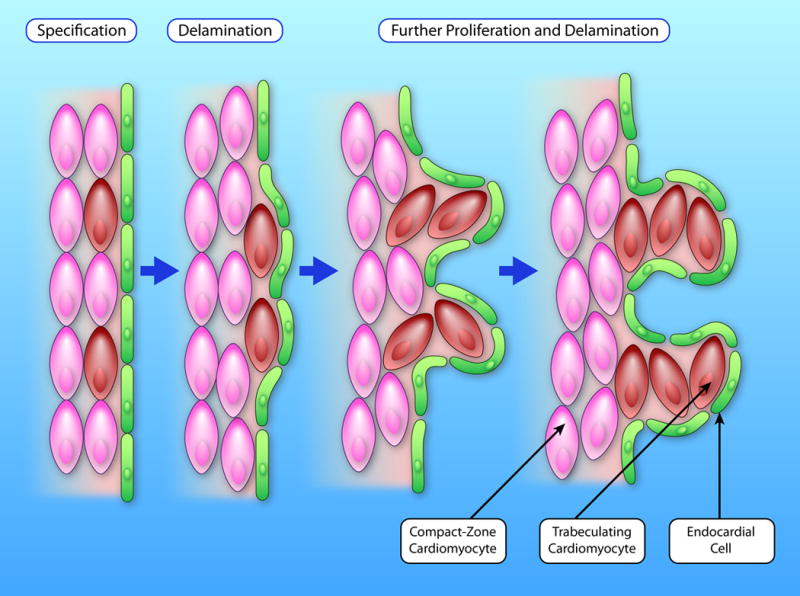
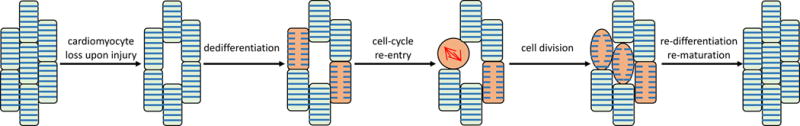
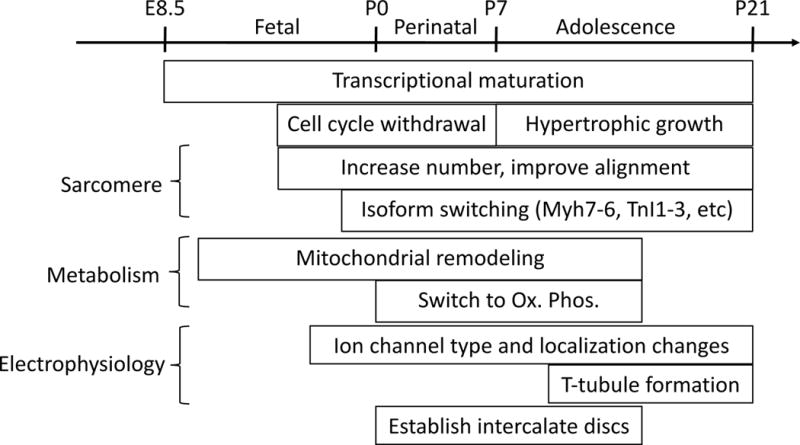
Palliative surgery for congenital heart disease has allowed patients with previously lethal heart malformations to survive and, in most cases, to thrive. However, these procedures often place pressure and volume loads on the heart, and over time, these chronic loads can cause heart failure. Current therapeutic options for initial surgery and chronic heart failure that results from failed palliation are limited, in part, by the mammalian heart's low inherent capacity to form new cardiomyocytes. Surmounting the heart regeneration barrier would transform the treatment of congenital, as well as acquired, heart disease and likewise would enable development of personalized, in vitro cardiac disease models. Although these remain distant goals, studies of heart development are illuminating the path forward and suggest unique opportunities for heart regeneration, particularly in fetal and neonatal periods. Here, we review major lessons from heart development that inform current and future studies directed at enhancing cardiac regeneration.
Keywords: cardiac development; cardiac regeneration; cardiomyocyte maturation; signaling pathways; transcriptional regulation.
© 2017 American Heart Association, Inc.
Conflict of interest statement
None
Figures
References
-
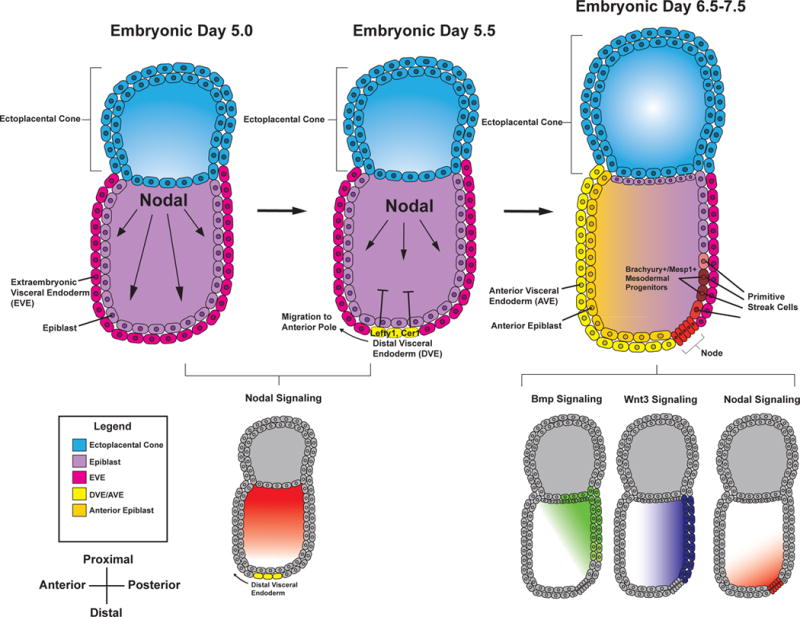
- Arnold SJ, Robertson EJ. Making a commitment: cell lineage allocation and axis patterning in the early mouse embryo. Nature Reviews Molecular Cell Biology. 2009;10:91–103. - PubMed
-
- Brennan J, Lu CC, Norris DP, Rodriguez TA. Nodal signalling in the epiblast patterns the early mouse embryo. Nodal signalling in the epiblast patterns the early mouse embryo. 2001 - PubMed
-
- Waldrip WR, Bikoff EK, Hoodless PA, Wrana JL, Robertson EJ. Smad2 signaling in extraembryonic tissues determines anterior-posterior polarity of the early mouse embryo. Cell. 1998;92:797–808. - PubMed
-
- Perea-Gomez A, Vella F, Shawlot W, Oulad-Abdelghani M, Chazaud C, Meno C, Pfister V, Chen L, Robertson E, Hamada H, Behringer RR, Ang S-L. Nodal Antagonists in the Anterior Visceral Endoderm Prevent the Formation of Multiple Primitive Streaks. Developmental Cell. 2002;3:745–756. - PubMed
-
- Conlon FL, Lyons KM, Takaesu N, Barth KS, Kispert A, Herrmann B, Robertson EJ. A primary requirement for nodal in the formation and maintenance of the primitive streak in the mouse. Development (Cambridge, England) 1994;120:1919–1928. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
