

Integration of optogenetics with complementary methodologies in systems neuroscience
- PMID: 28303019
- PMCID: PMC5708544
- DOI: 10.1038/nrn.2017.15
Integration of optogenetics with complementary methodologies in systems neuroscience
Abstract
Modern optogenetics can be tuned to evoke activity that corresponds to naturally occurring local or global activity in timing, magnitude or individual-cell patterning. This outcome has been facilitated not only by the development of core features of optogenetics over the past 10 years (microbial-opsin variants, opsin-targeting strategies and light-targeting devices) but also by the recent integration of optogenetics with complementary technologies, spanning electrophysiology, activity imaging and anatomical methods for structural and molecular analysis. This integrated approach now supports optogenetic identification of the native, necessary and sufficient causal underpinnings of physiology and behaviour on acute or chronic timescales and across cellular, circuit-level or brain-wide spatial scales.
Conflict of interest statement
The authors declare no competing interests.
Figures
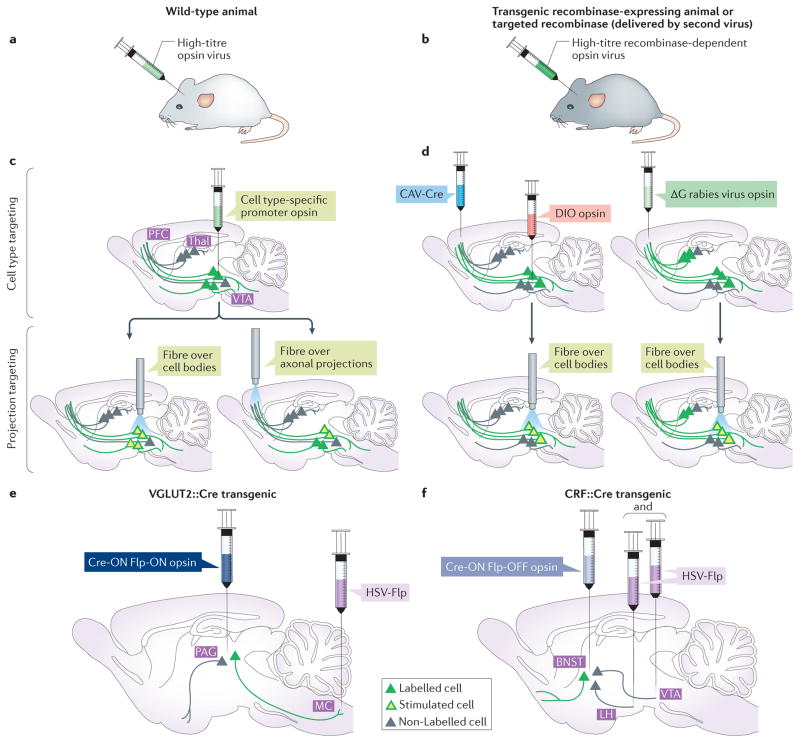
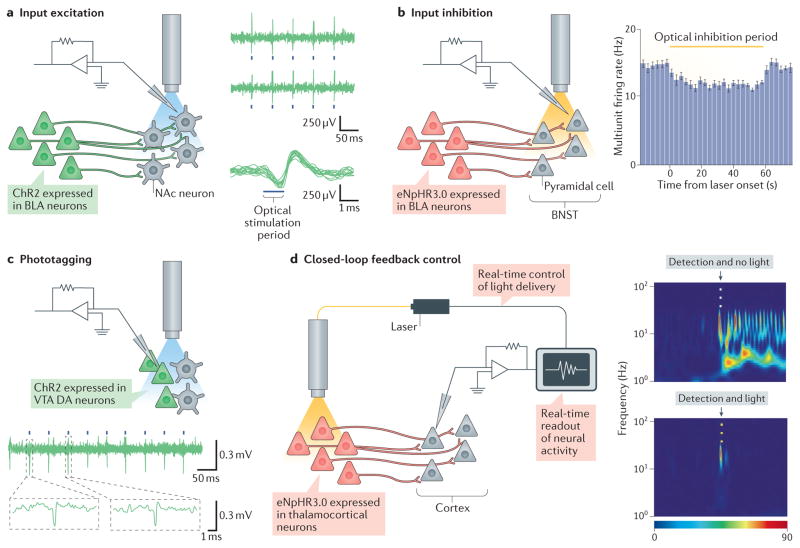
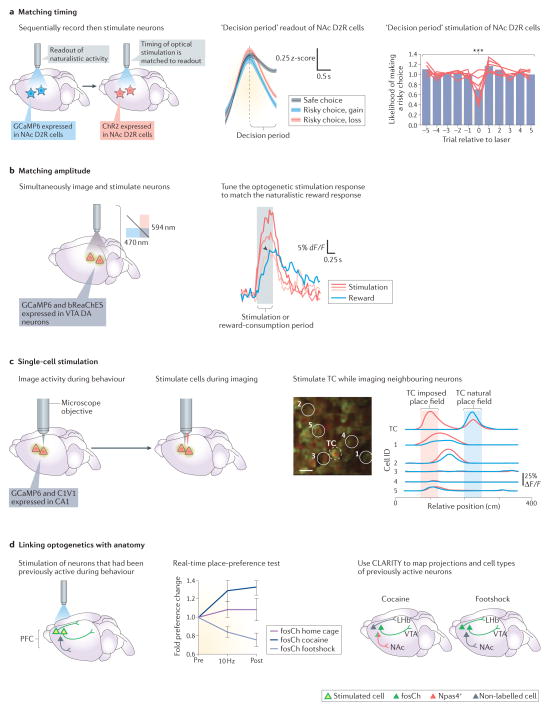
References
-
- Aravanis AM, et al. An optical neural interface: in vivo control of rodent motor cortex with integrated fiberoptic and optogenetic technology. J Neural Eng. 2007;4:S143–S156. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
