

Topography and Lateralized Effect of Acute Aromatase Inhibition on Auditory Processing in a Seasonal Songbird
- PMID: 28314822
- PMCID: PMC6596569
- DOI: 10.1523/JNEUROSCI.1961-16.2017
Topography and Lateralized Effect of Acute Aromatase Inhibition on Auditory Processing in a Seasonal Songbird
Abstract
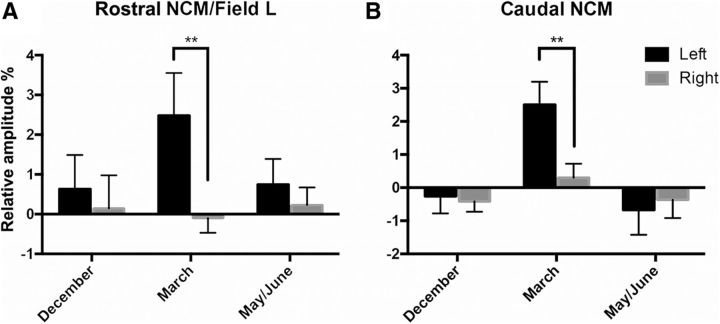
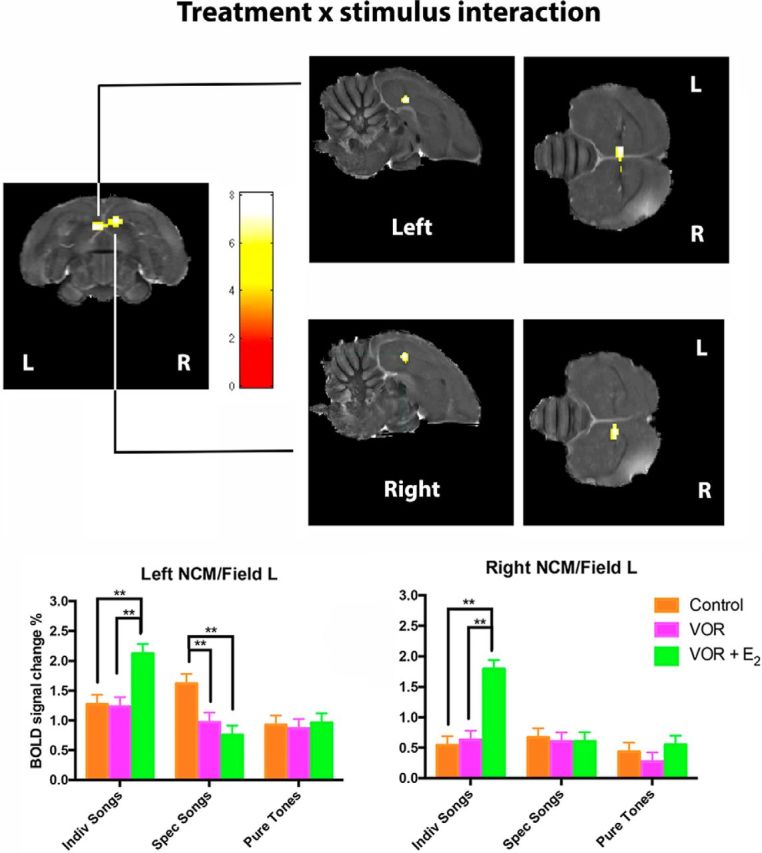
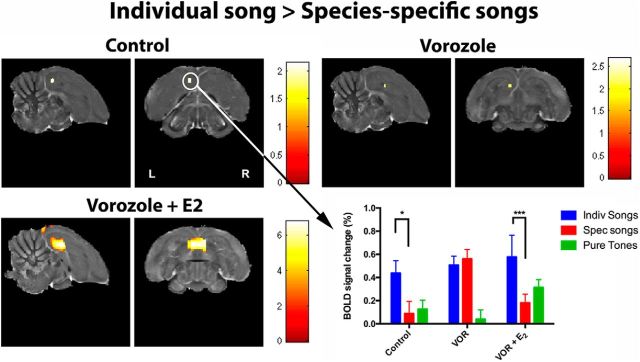
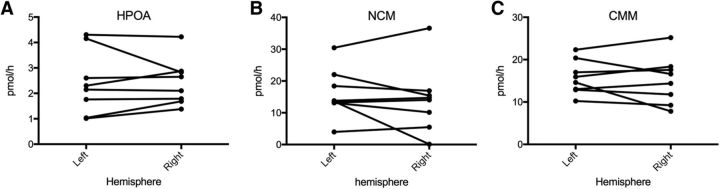
It is increasingly recognized that brain-derived estrogens (neuroestrogens) can regulate brain physiology and behavior much faster than what was previously known from the transcriptional action of estrogens on nuclear receptors. One of the best examples of such neuromodulation by neuroestrogens concerns the acute regulation of sensory coding by the auditory cortex as demonstrated by electrophysiological studies of selected neurons in zebra finches. Yet, the spatial extent of such modulation by neuroestrogens is not known. Using functional magnetic resonance imaging, we demonstrate here that acute estrogen depletion alters within minutes auditory processing in male European starlings. These effects are confined to very specific but large areas of the auditory cortex. They are also specifically lateralized to the left hemisphere. Interestingly, the modulation of auditory responses by estrogens was much larger (both in amplitude and in topography) in March than in December or May/June. This effect was presumably independent from changes in circulating testosterone concentrations since levels of the steroid were controlled by subcutaneous implants, thus suggesting actions related to other aspects of the seasonal cycle or photoperiodic manipulations. Finally, we also show that estrogen production specifically modulates selectivity for behaviorally relevant vocalizations in a specific part of the caudomedial nidopallium. These findings confirm and extend previous conclusions that had been obtained by electrophysiological techniques. This approach provides a new very powerful tool to investigate auditory responsiveness in songbirds and its fast modulation by sex steroids.SIGNIFICANCE STATEMENT Neuroestrogens can acutely modulate sensory processing in a manner similar to neuromodulators. We report that acute estrogen depletion rapidly disrupts auditory processing in large areas of the male starling brain. Effects were larger in March than in December or May/June, lateralized to the left hemisphere and specific to behaviorally relevant stimuli. These findings confirm and extend previous data that identified an acute regulation of auditory neurons in zebra finches by (1) delineating the extent of the brain region affected, (2) confirming its lateralization, and (3) demonstrating that a large part of the auditory brain regions are acutely affected by estrogens. These findings provide a very powerful tool to investigate auditory responsiveness in songbirds and its fast modulation by sex steroids.
Keywords: European starling; NCM; fMRI; lateralization; rapid estrogen action.
Copyright © 2017 the authors 0270-6474/17/374243-12$15.00/0.
Figures
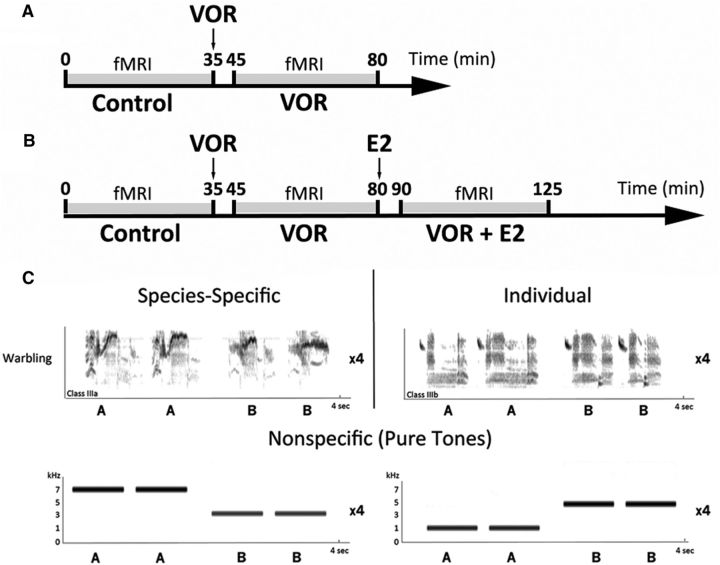
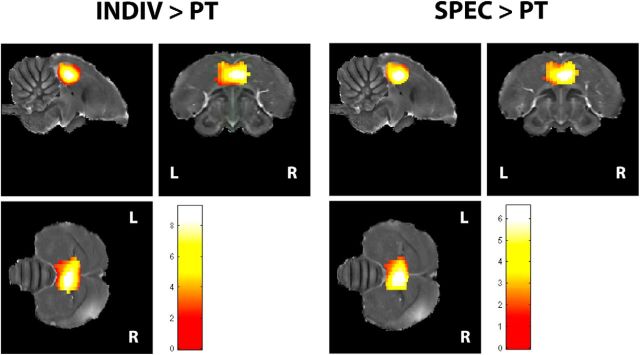
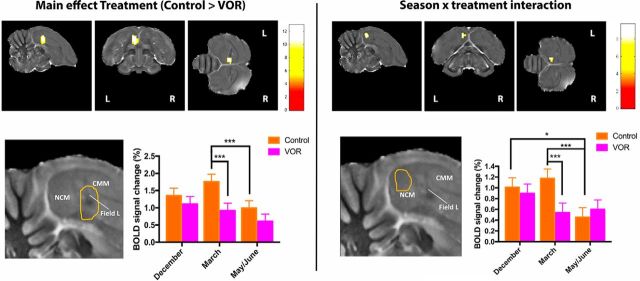
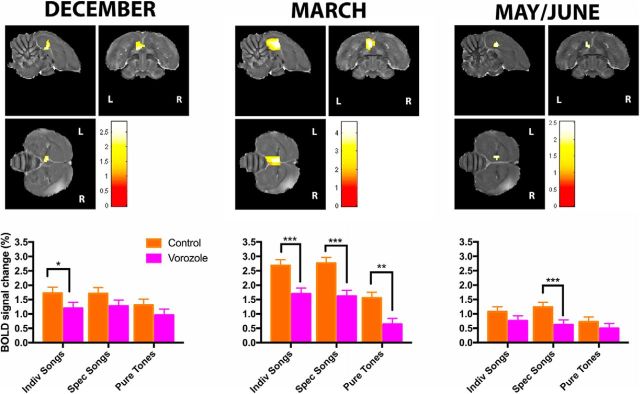
Comment in
-
Estradiol Modulates Hemispheric Lateralization of Auditory Evoked Neural Activity in Male European Starlings (Sturnus vulgaris).J Neurosci. 2017 Aug 16;37(33):7800-7802. doi: 10.1523/JNEUROSCI.1414-17.2017. J Neurosci. 2017. PMID: 28821600 Free PMC article. No abstract available.
References
-
- Apfelbeck B, Mortega K, Kiefer S, Kipper S, Vellema M, Villavicencio CP, Gahr M, Goymann W (2013) Associated and disassociated patterns in hormones, song, behavior and brain receptor expression between life-cycle stages in male black redstarts, Phoenicurus ochruros. Gen Comp Endocrinol 184:93–102. 10.1016/j.ygcen.2012.11.027 - DOI - PubMed
-
- Ball GF, Wingfield JC (1987) Changes in plasma levels of sex steroids in relation to multiple broodedness and nest site density in male starlings. PhysiolZool 60:191–196.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources