

Heme Gazing: Illuminating Eukaryotic Heme Trafficking, Dynamics, and Signaling with Fluorescent Heme Sensors
- PMID: 28316240
- PMCID: PMC5629415
- DOI: 10.1021/acs.biochem.7b00007
Heme Gazing: Illuminating Eukaryotic Heme Trafficking, Dynamics, and Signaling with Fluorescent Heme Sensors
Abstract
Heme (iron protoporphyrin IX) is an essential protein prosthetic group and signaling molecule required for most life on Earth. All heme-dependent processes require the dynamic and rapid mobilization of heme from sites of synthesis or uptake to hemoproteins present in virtually every subcellular compartment. The cytotoxicity and hydrophobicity of heme necessitate that heme mobilization be carefully controlled to mitigate the deleterious effects of this essential toxin. Indeed, a number of disorders, including certain cancers, cardiovascular diseases, and aging and age-related neurodegenerative diseases, are tied to defects in heme homeostasis. However, the molecules and mechanisms that mediate heme transport and trafficking, and the dynamics of these processes, are poorly understood. This is in large part due to the lack of physical tools for probing cellular heme. Herein, we discuss the recent development of fluorescent probes that can monitor and image kinetically labile heme with respect to its mobilization and role in signaling. In particular, we will highlight how heme gazing with these tools can uncover new heme trafficking factors upon being integrated with genetic screens and illuminate the concentration, subcellular distribution, and dynamics of labile heme in various physiological contexts. Altogether, the monitoring of labile heme, along with recent biochemical and cell biological studies demonstrating the reversible regulation of certain cellular processes by heme, is challenging us to reconceptualize heme from being a static cofactor buried in protein active sites to a dynamic and mobile signaling molecule.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
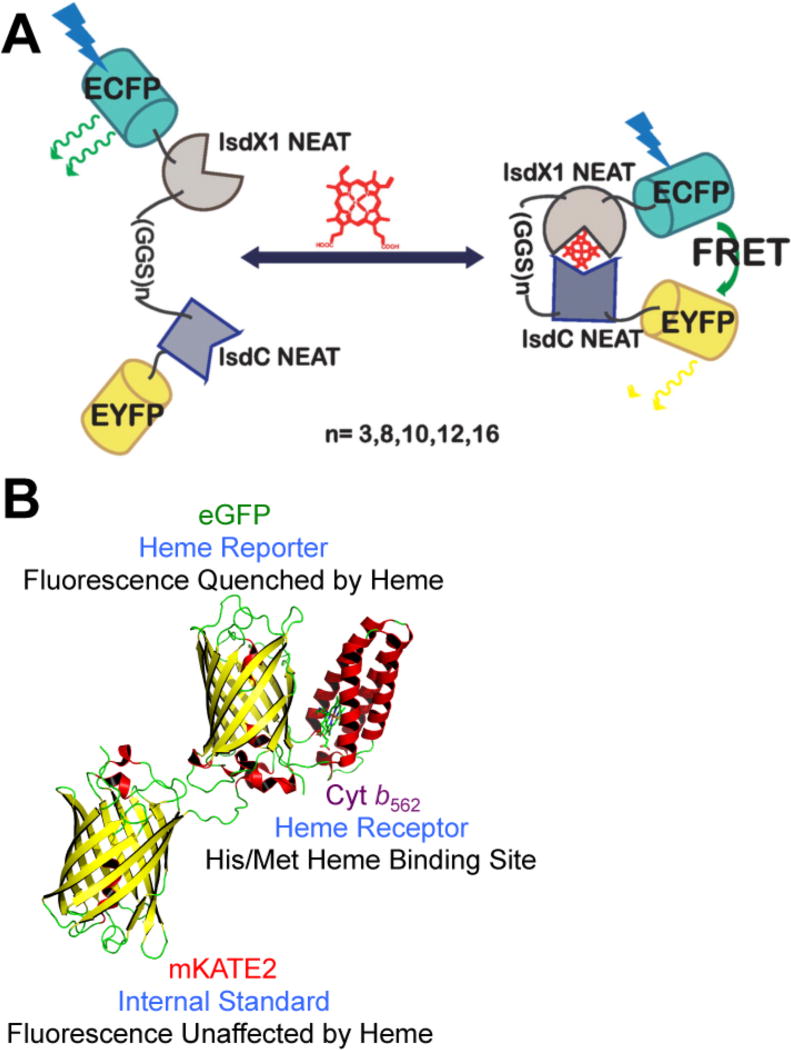
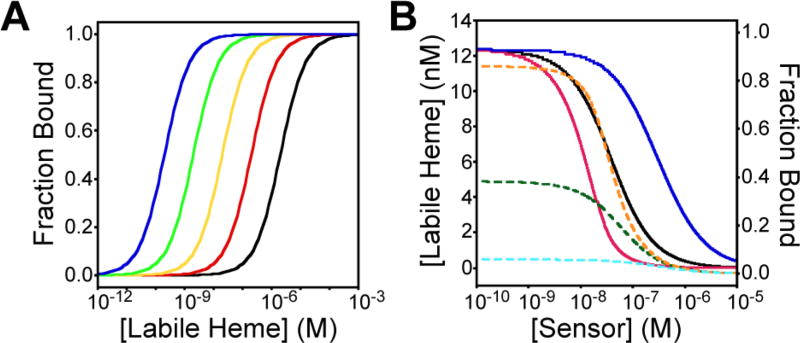
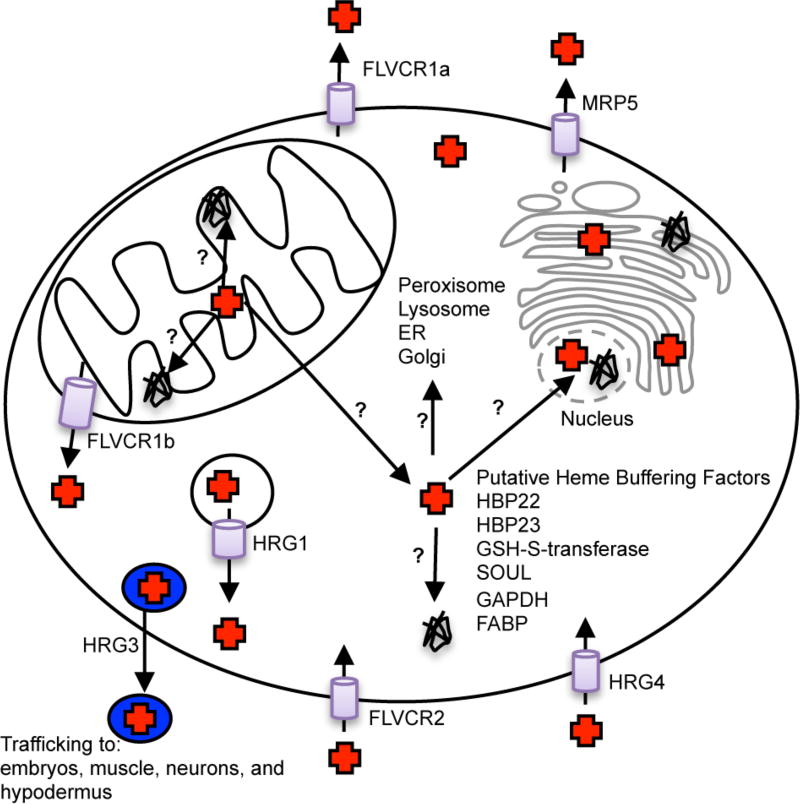
References
-
- Kumar S, Bandyopadhyay U. Free Heme Toxicity and Its Detoxification Systems in Human. Toxicol. Lett. 2005;157:175–188. - PubMed
-
- Sassa S. Why Heme Needs to Be Degraded to Iron, Biliverdin Ixalpha, and Carbon Monoxide? Antioxid. Redox Signaling. 2004;6:819–824. - PubMed
-
- Ponka P, Sheftel AD, English AM, Scott Bohle D, Garcia-Santos D. Do Mammalian Cells Really Need to Export and Import Heme? Trends Biochem. Sci 2017 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
