

Postmating Female Control: 20 Years of Cryptic Female Choice
- PMID: 28318651
- PMCID: PMC5511330
- DOI: 10.1016/j.tree.2017.02.010
Postmating Female Control: 20 Years of Cryptic Female Choice
Abstract
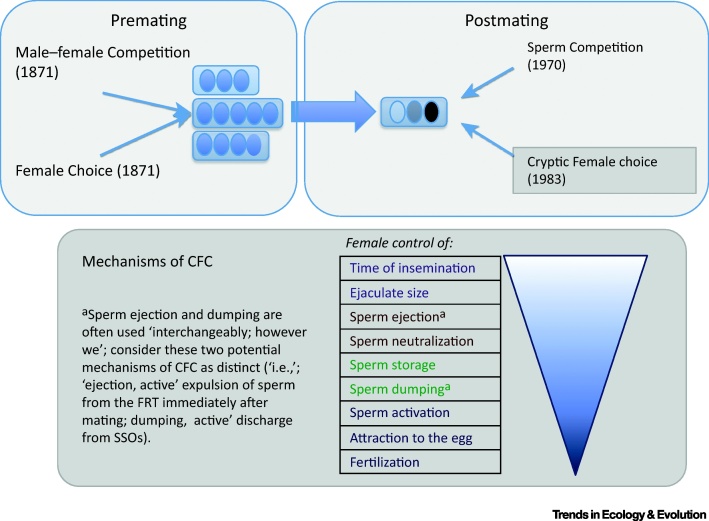
Cryptic female choice (CFC) represents postmating intersexual selection arising from female-driven mechanisms at or after mating that bias sperm use and impact male paternity share. Although biologists began to study CFC relatively late, largely spurred by Eberhard's book published 20 years ago, the field has grown rapidly since then. Here, we review empirical progress to show that numerous female processes offer potential for CFC, from mating through to fertilization, although seldom has CFC been clearly demonstrated. We then evaluate functional implications, and argue that, under some conditions, CFC might have repercussions for female fitness, sexual conflict, and intersexual coevolution, with ramifications for related evolutionary phenomena, such as speciation. We conclude by identifying directions for future research in this rapidly growing field.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures

Comment in
-
Cryptic Female Choice Revisited: A Response to Firman et al.Trends Ecol Evol. 2017 Nov;32(11):805-806. doi: 10.1016/j.tree.2017.08.009. Epub 2017 Sep 5. Trends Ecol Evol. 2017. PMID: 28886924 No abstract available.
-
Cryptic Female Choice: A General Phenomenon. A Reply to Eberhard.Trends Ecol Evol. 2017 Nov;32(11):807. doi: 10.1016/j.tree.2017.08.010. Epub 2017 Sep 7. Trends Ecol Evol. 2017. PMID: 28890125 No abstract available.
References
-
- Darwin C.R. John Murray; 1871. The Descent of Man and Selection in Relation to Sex.
-
- Parker G.A. Sperm competition and its evolutionary consequences in the insects. Biol. Rev. 1970;45:525–567.
-
- Childress D., Hartl D.L. Sperm preference in Drosophila melanogaster. Genetics. 1972;71:417–427. - PubMed
-
- Thornhill R. Cryptic female choice and its implications in the scorpionfly Harpobittacus nigriceps. Am. Nat. 1983;122:765–788.
-
- Eberhard W.G. Princeton University Press; 1996. Female Control: Sexual Selection by Cryptic Female Choice.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
