

Assembly of CNS Nodes of Ranvier in Myelinated Nerves Is Promoted by the Axon Cytoskeleton
- PMID: 28318976
- PMCID: PMC5387178
- DOI: 10.1016/j.cub.2017.01.025
Assembly of CNS Nodes of Ranvier in Myelinated Nerves Is Promoted by the Axon Cytoskeleton
Abstract
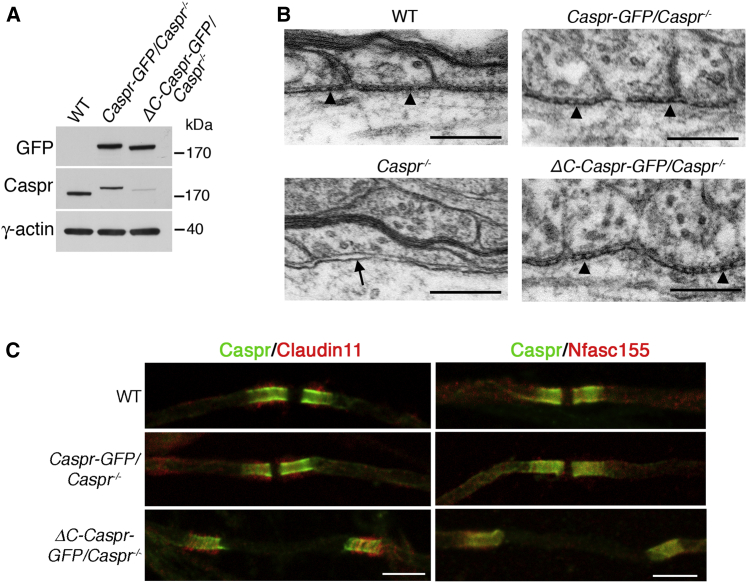
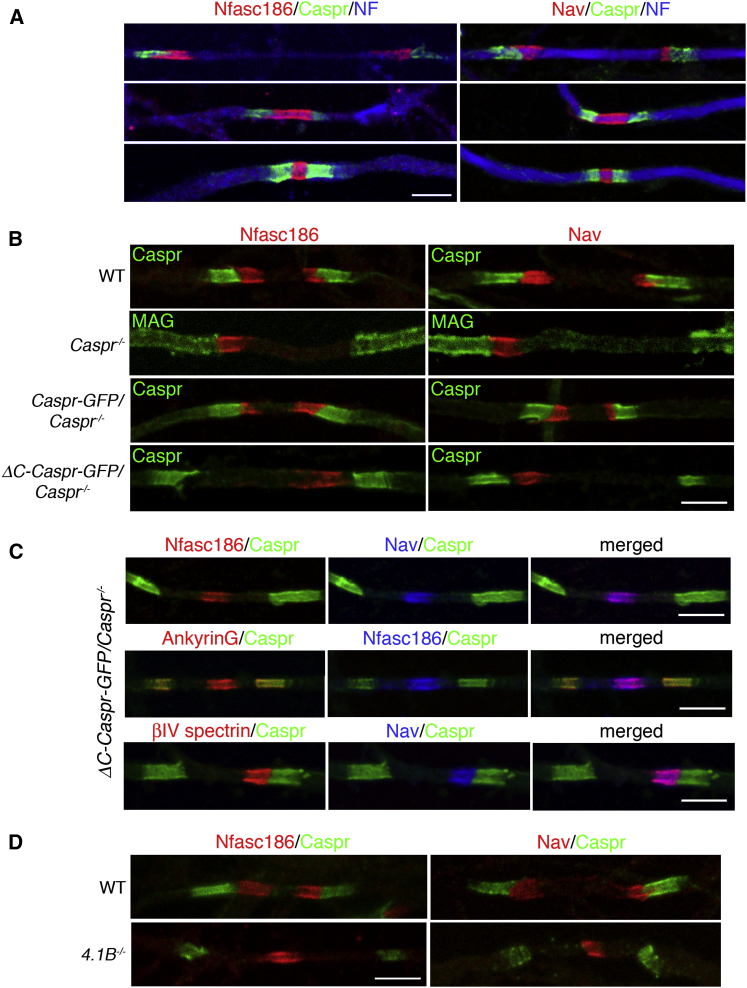
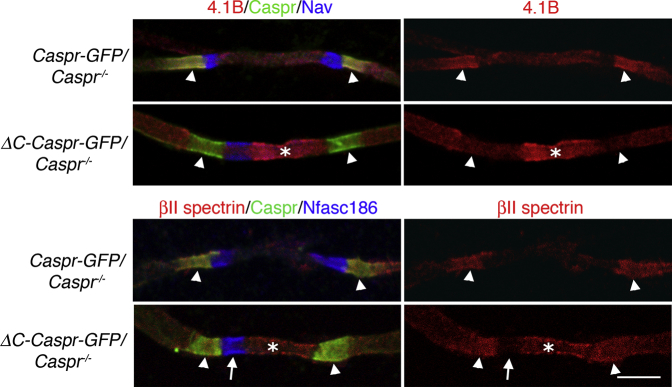
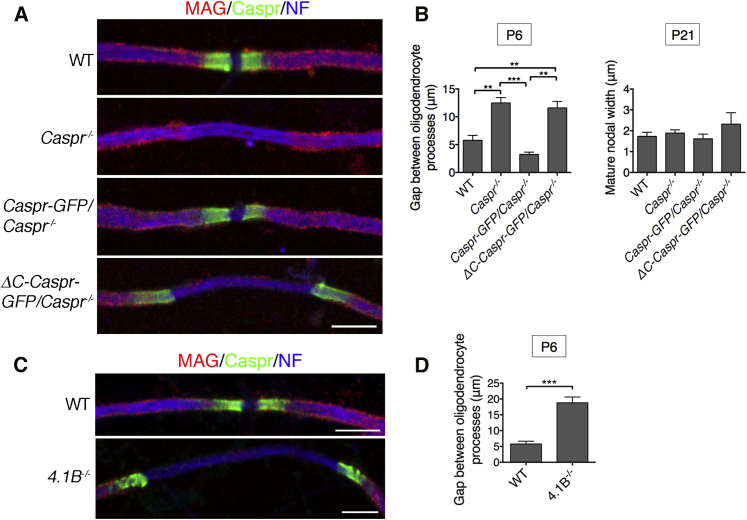
Nodes of Ranvier in the axons of myelinated neurons are exemplars of the specialized cell surface domains typical of polarized cells. They are rich in voltage-gated sodium channels (Nav) and thus underpin rapid nerve impulse conduction in the vertebrate nervous system [1]. Although nodal proteins cluster in response to myelination, how myelin-forming glia influence nodal assembly is poorly understood. An axoglial adhesion complex comprising glial Neurofascin155 and axonal Caspr/Contactin flanks mature nodes [2]. We have shown that assembly of this adhesion complex at the extremities of migrating oligodendroglial processes promotes process convergence along the axon during central nervous system (CNS) node assembly [3]. Here we show that anchorage of this axoglial complex to the axon cytoskeleton is essential for efficient CNS node formation. When anchorage is disrupted, both the adaptor Protein 4.1B and the cytoskeleton protein βII spectrin are mislocalized in the axon, and assembly of the node of Ranvier is significantly delayed. Nodal proteins and migrating oligodendroglial processes are no longer juxtaposed, and single detached nodal complexes replace the symmetrical heminodes found in both the CNS and peripheral nervous system (PNS) during development. We propose that axoglial adhesion complexes contribute to the formation of an interface between cytoskeletal elements enriched in Protein 4.1B and βII spectrin and those enriched in nodal ankyrinG and βIV spectrin. This clusters nascent nodal complexes at heminodes and promotes their timely coalescence to form the mature node of Ranvier. These data demonstrate a role for the axon cytoskeleton in the assembly of a critical neuronal domain, the node of Ranvier.
Keywords: CNS; myelination; node of Ranvier; paranodal axoglial junction.
Copyright © 2017 The Author(s). Published by Elsevier Ltd.. All rights reserved.
Figures
References
-
- Huxley A.F., Stämpfli R. Evidence for saltatory conduction in peripheral myelinated nerve fibres. J. Physiol. 1949;108:315–339. - PubMed
-
- Sherman D.L., Tait S., Melrose S., Johnson R., Zonta B., Court F.A., Macklin W.B., Meek S., Smith A.J., Cottrell D.F., Brophy P.J. Neurofascins are required to establish axonal domains for saltatory conduction. Neuron. 2005;48:737–742. - PubMed
-
- Bhat M.A., Rios J.C., Lu Y., Garcia-Fresco G.P., Ching W., St Martin M., Li J., Einheber S., Chesler M., Rosenbluth J. Axon-glia interactions and the domain organization of myelinated axons requires neurexin IV/Caspr/Paranodin. Neuron. 2001;30:369–383. - PubMed
-
- Boyle M.E., Berglund E.O., Murai K.K., Weber L., Peles E., Ranscht B. Contactin orchestrates assembly of the septate-like junctions at the paranode in myelinated peripheral nerve. Neuron. 2001;30:385–397. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
