

Calcium channel modulation as a target in chronic pain control
- PMID: 28320042
- PMCID: PMC5980588
- DOI: 10.1111/bph.13789
Calcium channel modulation as a target in chronic pain control
Abstract
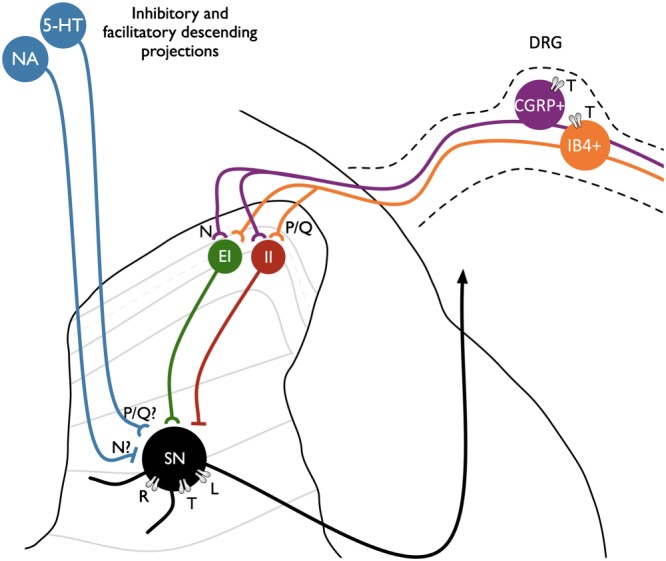
Neuropathic pain remains poorly treated for large numbers of patients, and little progress has been made in developing novel classes of analgesics. To redress this issue, ziconotide (Prialt™) was developed and approved as a first-in-class synthetic version of ω-conotoxin MVIIA, a peptide blocker of Cav 2.2 channels. Unfortunately, the impracticalities of intrathecal delivery, low therapeutic index and severe neurological side effects associated with ziconotide have restricted its use to exceptional circumstances. Ziconotide exhibits no state or use-dependent block of Cav 2.2 channels; activation state-dependent blockers were hypothesized to circumvent the side effects of state-independent blockers by selectively targeting high-frequency firing of nociceptive neurones in chronic pain states, thus alleviating aberrant pain but not affecting normal sensory transduction. Unfortunately, numerous drugs, including state-dependent calcium channel blockers, have displayed efficacy in preclinical models but have subsequently been disappointing in clinical trials. In recent years, it has become more widely acknowledged that trans-aetiological sensory profiles exist amongst chronic pain patients and may indicate similar underlying mechanisms and drug sensitivities. Heterogeneity amongst patients, a reliance on stimulus-evoked endpoints in preclinical studies and a failure to utilize translatable endpoints, all are likely to have contributed to negative clinical trial results. We provide an overview of how electrophysiological and operant-based assays provide insight into sensory and affective aspects of pain in animal models and how these may relate to chronic pain patients in order to improve the bench-to-bedside translation of calcium channel modulators.
Linked articles: This article is part of a themed section on Recent Advances in Targeting Ion Channels to Treat Chronic Pain. To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v175.12/issuetoc.
© 2017 The Authors. British Journal of Pharmacology published by John Wiley & Sons Ltd on behalf of British Pharmacological Society.
Figures


References
-
- Abbadie C, McManus OB, Sun SY, Bugianesi RM, Dai G, Haedo RJ et al. (2010). Analgesic effects of a substituted N‐triazole oxindole (TROX‐1), a state‐dependent, voltage‐gated calcium channel 2 blocker. J Pharmacol Exp Ther 334: 545–555. - PubMed
-
- Andrews N, Loomis S, Blake R, Ferrigan L, Singh L, McKnight AT (2001). Effect of gabapentin‐like compounds on development and maintenance of morphine‐induced conditioned place preference. Psychopharmacology (Berl) 157: 381–387. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
