

Dexpramipexole improves bioenergetics and outcome in experimental stroke
- PMID: 28320070
- PMCID: PMC5758384
- DOI: 10.1111/bph.13790
Dexpramipexole improves bioenergetics and outcome in experimental stroke
Abstract
Background and purpose: Dexpramipexole, a drug recently tested in patients with amyotrophic lateral sclerosis (ALS,) is able to bind F1Fo ATP synthase and increase mitochondrial ATP production. Here, we have investigated its effects on experimental ischaemic brain injury.
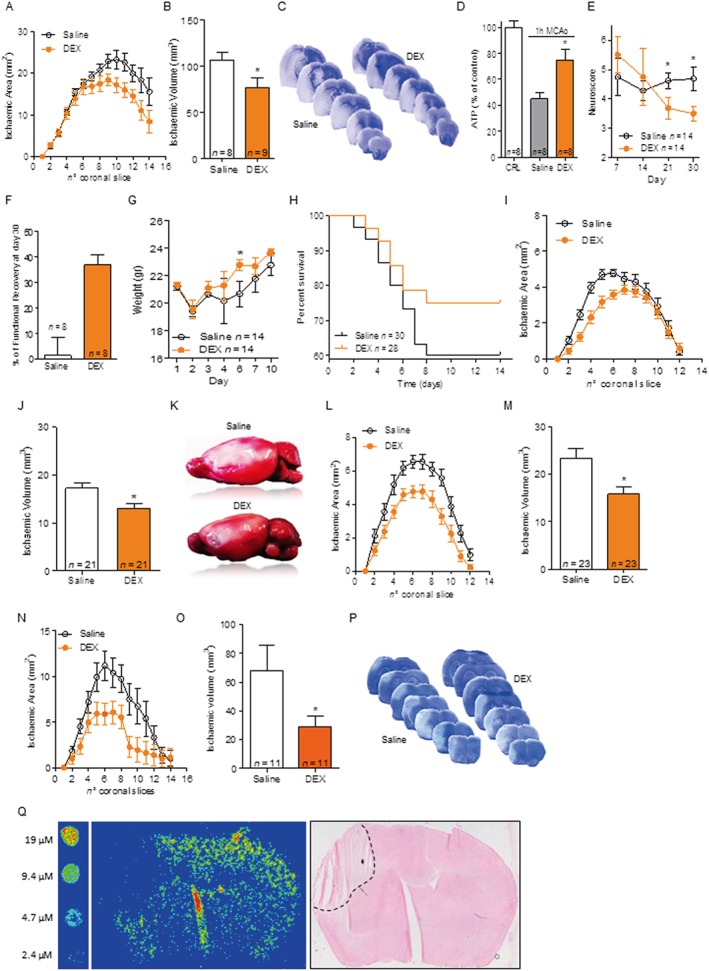
Experimental approach: The effects of dexpramipexole on bioenergetics, Ca2+ fluxes, electrophysiological functions and death were evaluated in primary neural cultures and hippocampal slices exposed to oxygen-glucose deprivation (OGD). Effects on infarct volumes and neurological functions were also evaluated in mice following proximal or distal middle cerebral artery occlusion (MCAo). Distribution of dexpramipexole within the ischaemic brain was evaluated by means of mass spectrometry imaging.
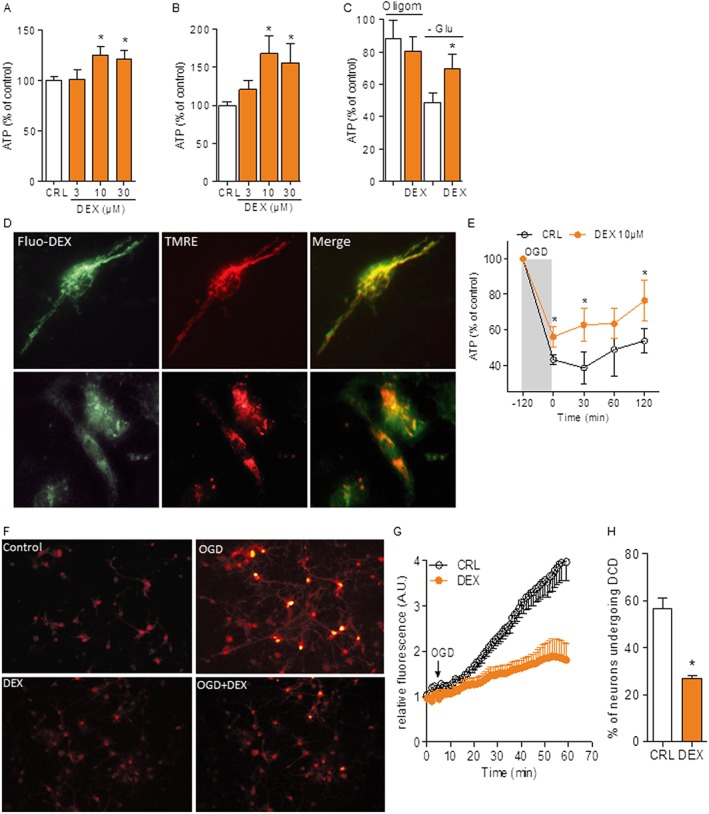
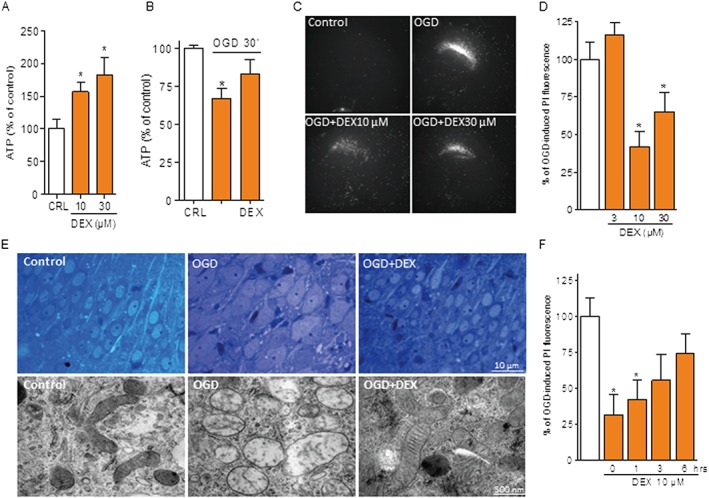
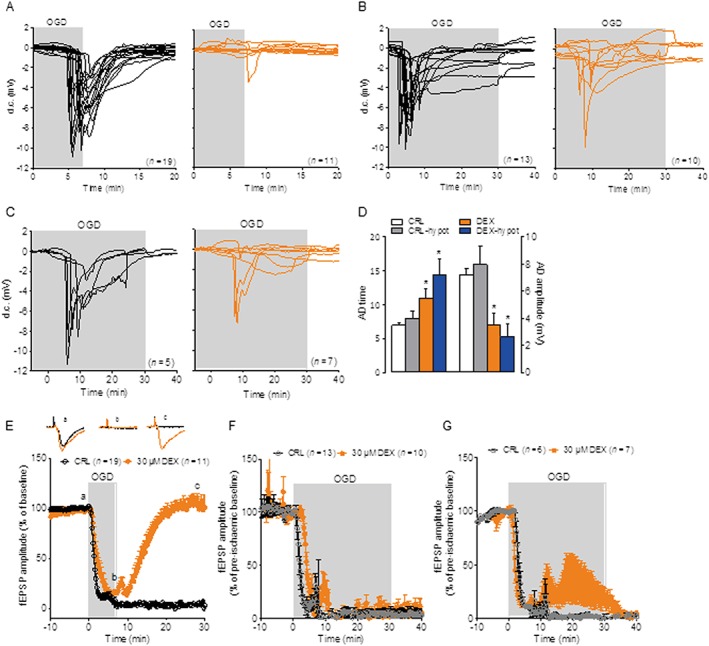
Key results: Dexpramipexole increased mitochondrial ATP production in cultured neurons or glia and reduces energy failure, prevents intracellular Ca2+ overload and affords cytoprotection when cultures are exposed to OGD. This compound also counteracted ATP depletion, mitochondrial swelling, anoxic depolarization, loss of synaptic activity and neuronal death in hippocampal slices subjected to OGD. Post-ischaemic treatment with dexpramipexole, at doses consistent with those already used in ALS patients, reduced brain infarct size and ameliorated neuroscore in mice subjected to transient or permanent MCAo. Notably, the concentrations of dexpramipexole reached within the ischaemic penumbra equalled those found neuroprotective in vitro.
Conclusion and implications: Dexpramipexole, a compound able to increase mitochondrial F1Fo ATP-synthase activity reduced ischaemic brain injury. These findings, together with the excellent brain penetration and favourable safety profile in humans, make dexpramipexole a drug with realistic translational potential for the treatment of stroke.
Linked articles: This article is part of a themed section on Inventing New Therapies Without Reinventing the Wheel: The Power of Drug Repurposing. To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v175.2/issuetoc.
© 2017 The British Pharmacological Society.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
