

Mechanisms to Mitigate the Trade-Off between Growth and Defense
- PMID: 28320784
- PMCID: PMC5435432
- DOI: 10.1105/tpc.16.00931
Mechanisms to Mitigate the Trade-Off between Growth and Defense
Abstract
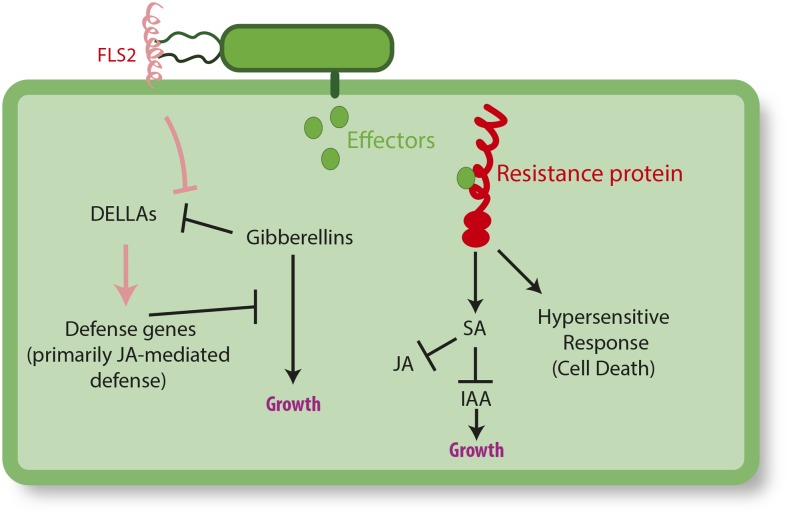
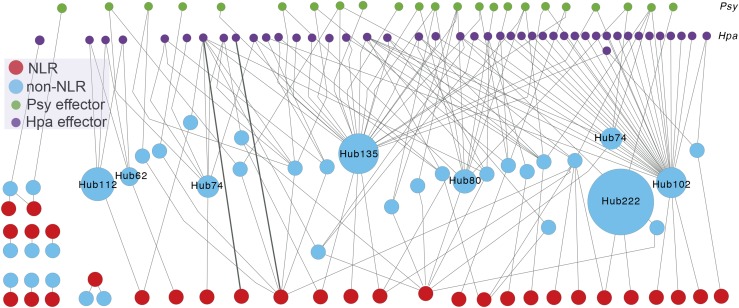
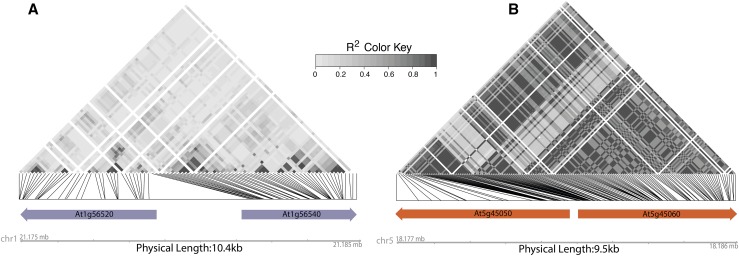
Plants have evolved an array of defenses against pathogens. However, mounting a defense response frequently comes with the cost of a reduction in growth and reproduction, carrying critical implications for natural and agricultural populations. This review focuses on how costs are generated and whether and how they can be mitigated. Most well-characterized growth-defense trade-offs stem from antagonistic crosstalk among hormones rather than an identified metabolic expenditure. A primary way plants mitigate such costs is through restricted expression of resistance; this can be achieved through inducible expression of defense genes or by the concentration of defense to particular times or tissues. Defense pathways can be primed for more effective induction, and primed states can be transmitted to offspring. We examine the resistance (R) genes as a case study of how the toll of defense can be generated and ameliorated. The fine-scale regulation of R genes is critical to alleviate the burden of their expression, and the genomic organization of R genes into coregulatory modules reduces costs. Plants can also recruit protection from other species. Exciting new evidence indicates that a plant's genotype influences the microbiome composition, lending credence to the hypothesis that plants shape their microbiome to enhance defense.
© 2017 American Society of Plant Biologists. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
