

Molecular mechanism of dynein recruitment to kinetochores by the Rod-Zw10-Zwilch complex and Spindly
- PMID: 28320824
- PMCID: PMC5379953
- DOI: 10.1083/jcb.201610108
Molecular mechanism of dynein recruitment to kinetochores by the Rod-Zw10-Zwilch complex and Spindly
Abstract
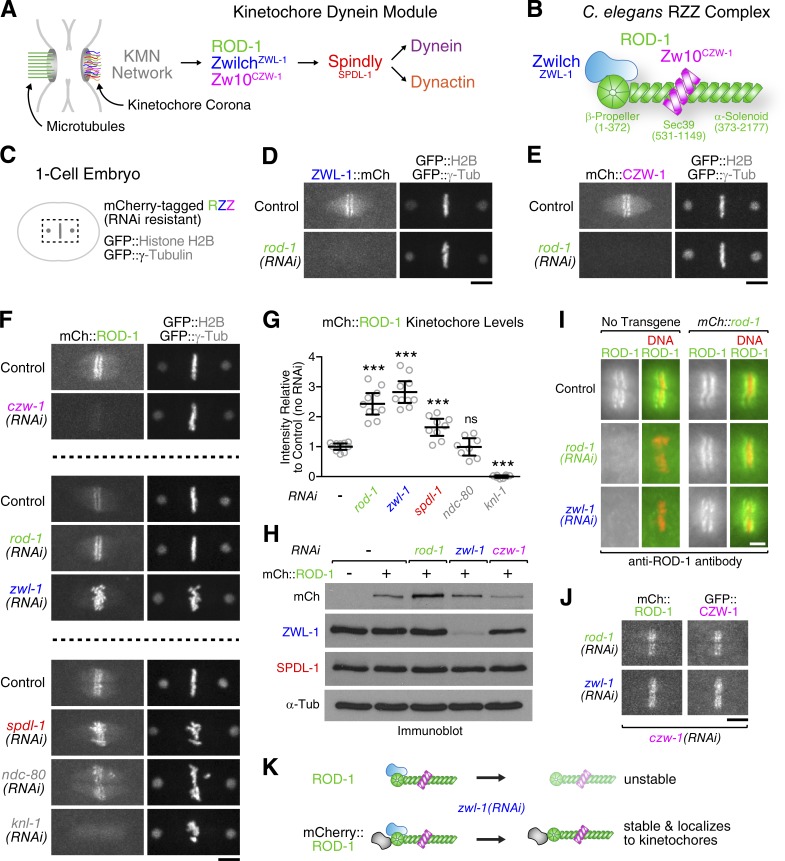
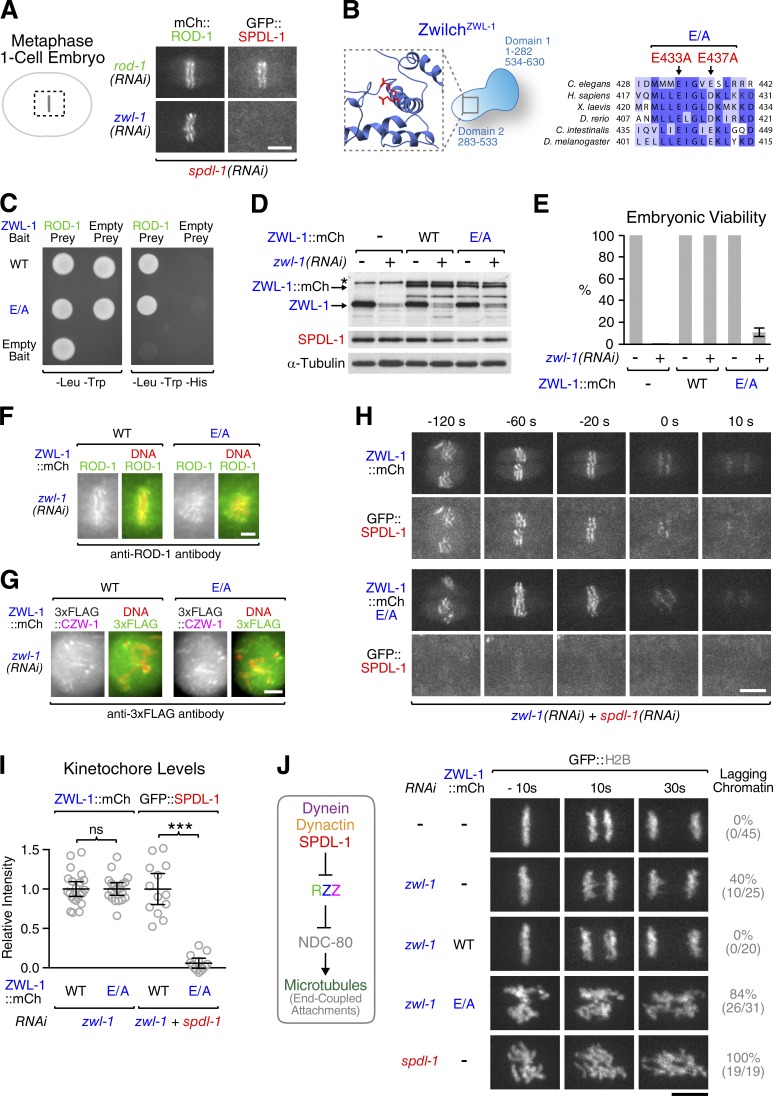
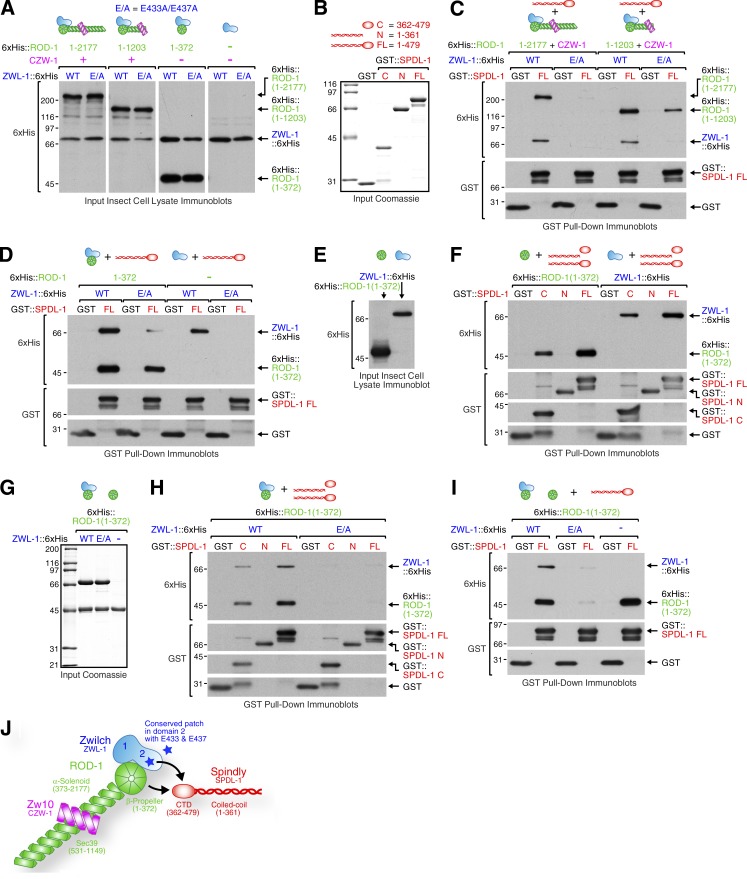
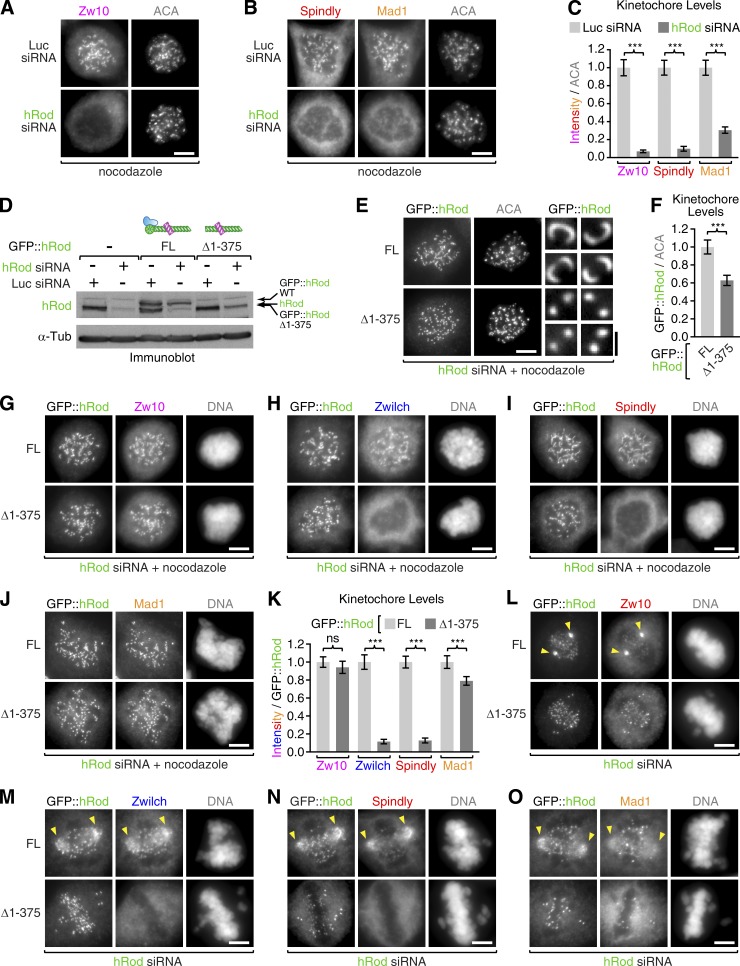
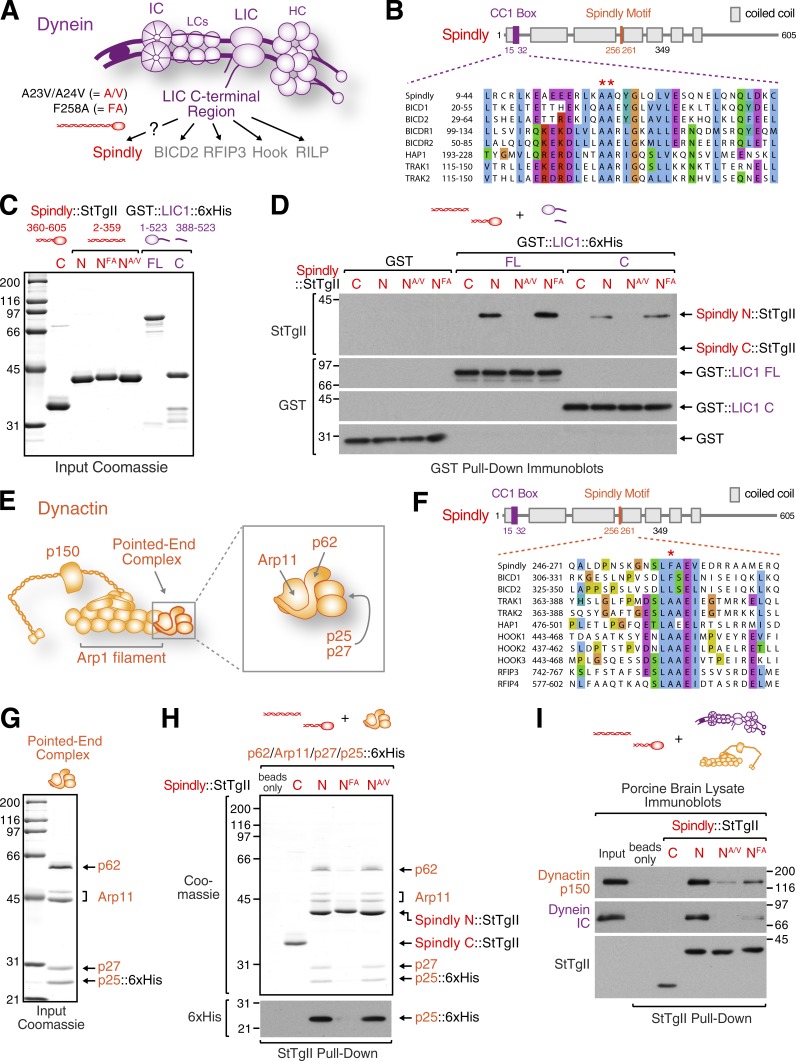
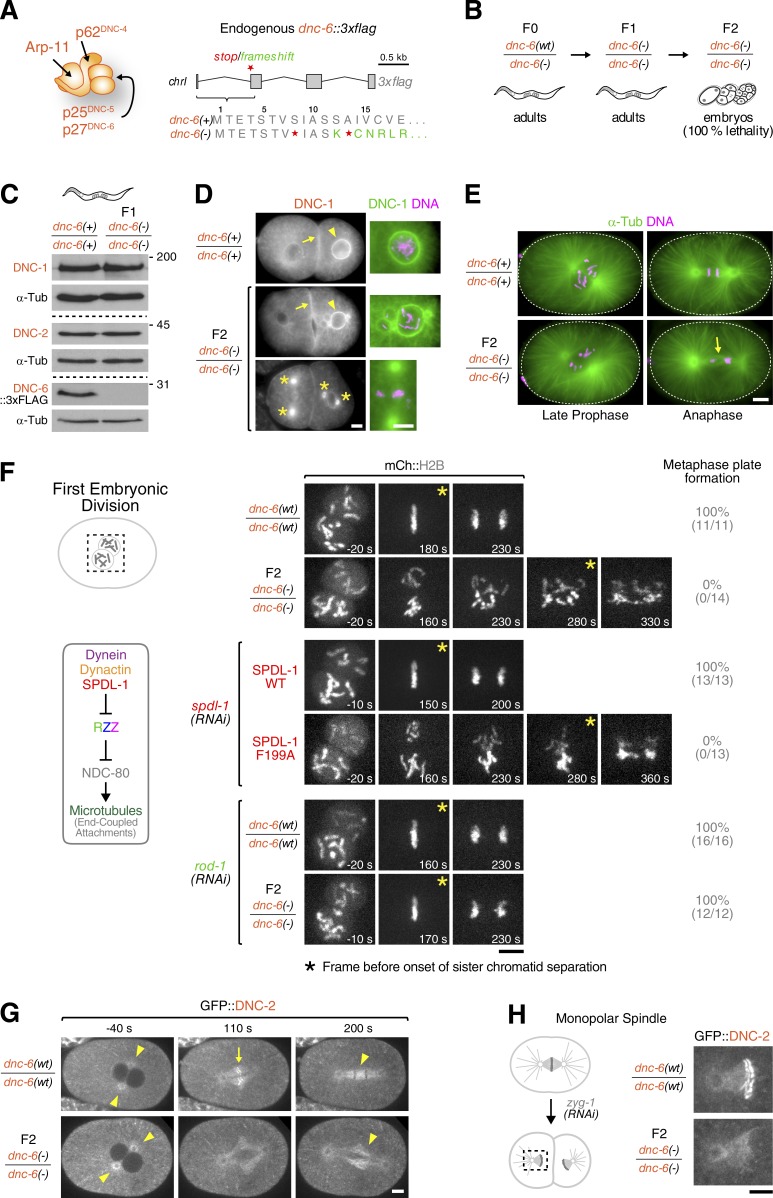
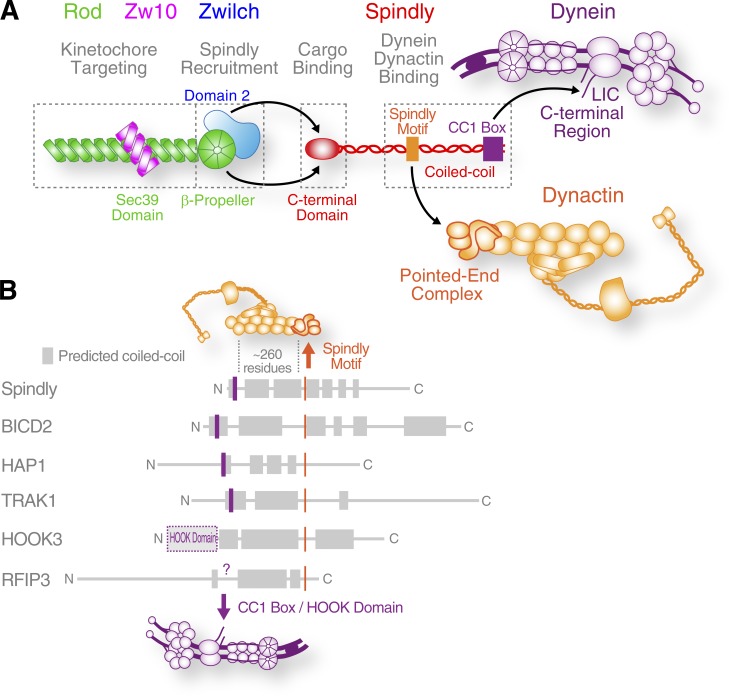
The molecular motor dynein concentrates at the kinetochore region of mitotic chromosomes in animals to accelerate spindle microtubule capture and to control spindle checkpoint signaling. In this study, we describe the molecular mechanism used by the Rod-Zw10-Zwilch complex and the adaptor Spindly to recruit dynein to kinetochores in Caenorhabditis elegans embryos and human cells. We show that Rod's N-terminal β-propeller and the associated Zwilch subunit bind Spindly's C-terminal domain, and we identify a specific Zwilch mutant that abrogates Spindly and dynein recruitment in vivo and Spindly binding to a Rod β-propeller-Zwilch complex in vitro. Spindly's N-terminal coiled-coil uses distinct motifs to bind dynein light intermediate chain and the pointed-end complex of dynactin. Mutations in these motifs inhibit assembly of a dynein-dynactin-Spindly complex, and a null mutant of the dynactin pointed-end subunit p27 prevents kinetochore recruitment of dynein-dynactin without affecting other mitotic functions of the motor. Conservation of Spindly-like motifs in adaptors involved in intracellular transport suggests a common mechanism for linking dynein to cargo.
Keywords: mitosis; kinetochore; dynein/dynactin; BICD2; Spindly; RZZ complex.
© 2017 Gama et al.
Figures
References
-
- Bieniossek C., Richmond T.J., and Berger I.. 2008. MultiBac: multigene baculovirus-based eukaryotic protein complex production. Curr. Protoc. Protein Sci. 5:Unit 5.20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
