

Neuroendocrine androgen action is a key extraovarian mediator in the development of polycystic ovary syndrome
- PMID: 28320971
- PMCID: PMC5402450
- DOI: 10.1073/pnas.1616467114
Neuroendocrine androgen action is a key extraovarian mediator in the development of polycystic ovary syndrome
Abstract
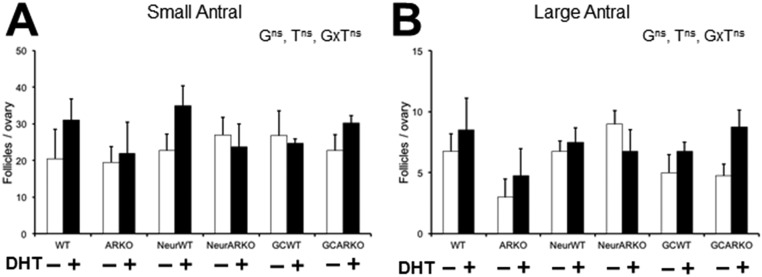
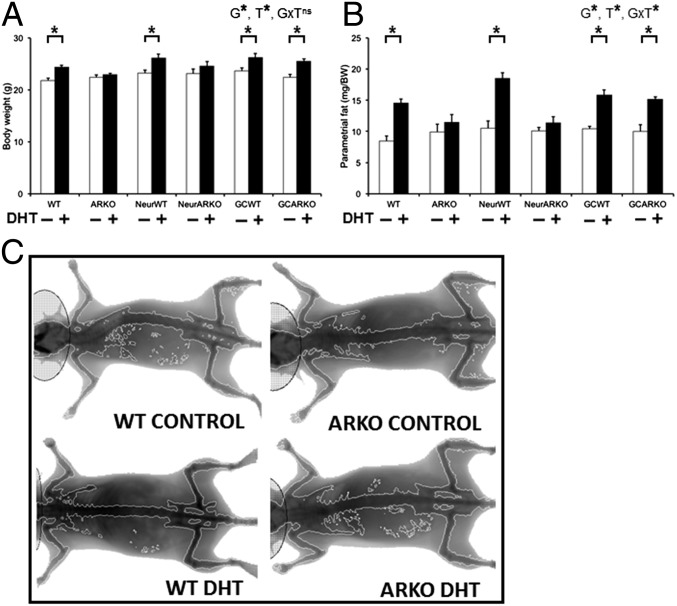
Polycystic ovary syndrome (PCOS) is a complex hormonal disorder characterized by reproductive, endocrine, and metabolic abnormalities. As the origins of PCOS remain unknown, mechanism-based treatments are not feasible and current management relies on treatment of symptoms. Hyperandrogenism is the most consistent PCOS characteristic; however, it is unclear whether androgen excess, which is treatable, is a cause or a consequence of PCOS. As androgens mediate their actions via the androgen receptor (AR), we combined a mouse model of dihydrotestosterone (DHT)-induced PCOS with global and cell-specific AR-resistant (ARKO) mice to investigate the locus of androgen actions that mediate the development of the PCOS phenotype. Global loss of the AR reveals that AR signaling is required for all DHT-induced features of PCOS. Neuron-specific AR signaling was required for the development of dysfunctional ovulation, classic polycystic ovaries, reduced large antral follicle health, and several metabolic traits including obesity and dyslipidemia. In addition, ovariectomized ARKO hosts with wild-type ovary transplants displayed normal estrous cycles and corpora lutea, despite DHT treatment, implying extraovarian and not intraovarian AR actions are key loci of androgen action in generating the PCOS phenotype. These findings provide strong evidence that neuroendocrine genomic AR signaling is an important extraovarian mediator in the development of PCOS traits. Thus, targeting AR-driven mechanisms that initiate PCOS is a promising strategy for the development of novel treatments for PCOS.
Keywords: PCOS; androgen; animal model; neuroendocrine.
Conflict of interest statement
The authors declare no conflict of interest.
Figures









Comment in
-
Neuronal androgen receptor: Molecular gateway to polycystic ovary syndrome?Proc Natl Acad Sci U S A. 2017 Apr 18;114(16):4045-4047. doi: 10.1073/pnas.1703436114. Epub 2017 Apr 4. Proc Natl Acad Sci U S A. 2017. PMID: 28377515 Free PMC article. No abstract available.
-
PCOS: Animal models for PCOS - not the real thing.Nat Rev Endocrinol. 2017 Jul;13(7):382-384. doi: 10.1038/nrendo.2017.57. Epub 2017 May 5. Nat Rev Endocrinol. 2017. PMID: 28474686 No abstract available.
References
-
- Fauser BC, et al. Consensus on women’s health aspects of polycystic ovary syndrome (PCOS): The Amsterdam ESHRE/ASRM-Sponsored 3rd PCOS Consensus Workshop Group. Fertil Steril. 2012;97(1):28–38.e25. - PubMed
-
- Gorry A, White DM, Franks S. Infertility in polycystic ovary syndrome: Focus on low-dose gonadotropin treatment. Endocrine. 2006;30(1):27–33. - PubMed
-
- Shorakae S, Boyle J, Teede H. Polycystic ovary syndrome: A common hormonal condition with major metabolic sequelae that physicians should know about. Intern Med J. 2014;44(8):720–726. - PubMed
-
- Nisenblat V, Norman RJ. Androgens and polycystic ovary syndrome. Curr Opin Endocrinol Diabetes Obes. 2009;16(3):224–231. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
