

Small RNA-mediated repair of UV-induced DNA lesions by the DNA DAMAGE-BINDING PROTEIN 2 and ARGONAUTE 1
- PMID: 28325872
- PMCID: PMC5389294
- DOI: 10.1073/pnas.1618834114
Small RNA-mediated repair of UV-induced DNA lesions by the DNA DAMAGE-BINDING PROTEIN 2 and ARGONAUTE 1
Abstract
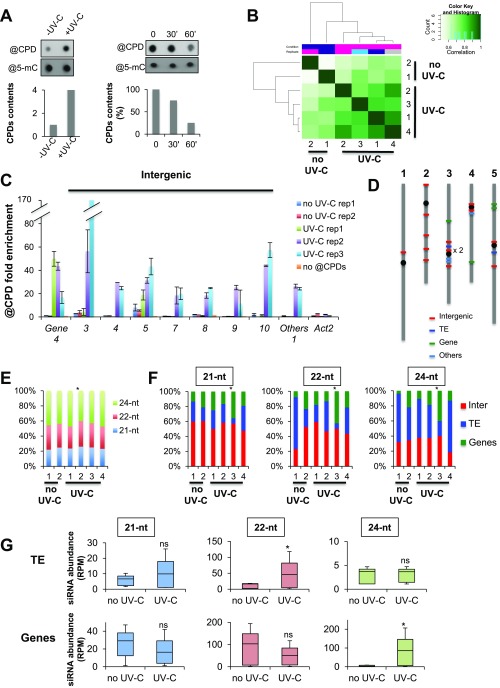
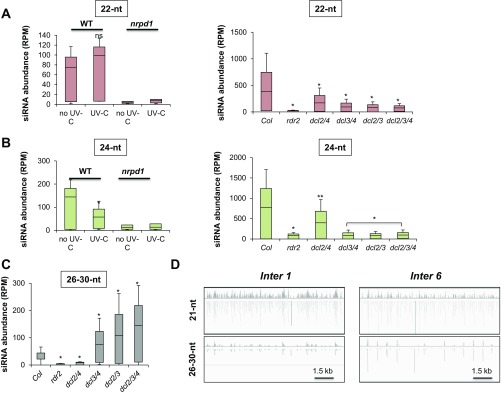
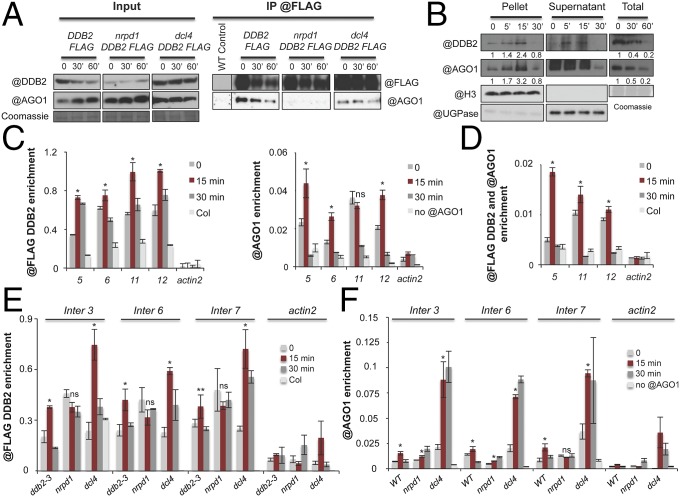
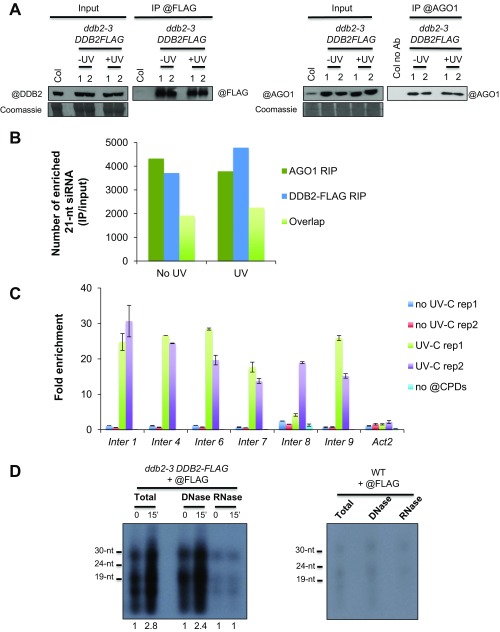
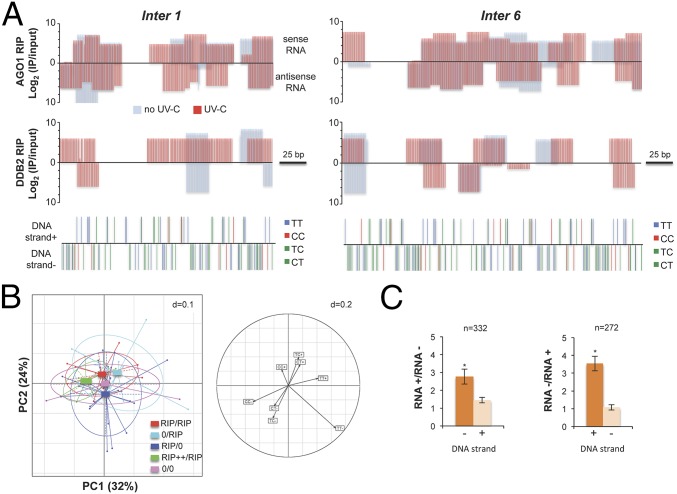
As photosynthetic organisms, plants need to prevent irreversible UV-induced DNA lesions. Through an unbiased, genome-wide approach, we have uncovered a previously unrecognized interplay between Global Genome Repair and small interfering RNAs (siRNAs) in the recognition of DNA photoproducts, prevalently in intergenic regions. Genetic and biochemical approaches indicate that, upon UV irradiation, the DNA DAMAGE-BINDING PROTEIN 2 (DDB2) and ARGONAUTE 1 (AGO1) of Arabidopsis thaliana form a chromatin-bound complex together with 21-nt siRNAs, which likely facilitates recognition of DNA damages in an RNA/DNA complementary strand-specific manner. The biogenesis of photoproduct-associated siRNAs involves the noncanonical, concerted action of RNA POLYMERASE IV, RNA-DEPENDENT RNA POLYMERASE-2, and DICER-LIKE-4. Furthermore, the chromatin association/dissociation of the DDB2-AGO1 complex is under the control of siRNA abundance and DNA damage signaling. These findings reveal unexpected nuclear functions for DCL4 and AGO1, and shed light on the interplay between small RNAs and DNA repair recognition factors at damaged sites.
Keywords: Arabidopsis; DNA photolesions; DNA repair; small RNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








References
-
- Schreier WJ, Gilch P, Zinth W. Early events of DNA photodamage. Annu Rev Phys Chem. 2015;66:497–519. - PubMed
-
- Sancar A. Photolyase and cryptochrome blue-light photoreceptors. Adv Protein Chem. 2004;69:73–100. - PubMed
-
- Chu G, Chang E. Xeroderma pigmentosum group E cells lack a nuclear factor that binds to damaged DNA. Science. 1988;242:564–567. - PubMed
-
- Nichols AF, et al. Human damage-specific DNA-binding protein p48. Characterization of XPE mutations and regulation following UV irradiation. J Biol Chem. 2000;275:21422–21428. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
