

Ion and inhibitor binding of the double-ring ion selectivity filter of the mitochondrial calcium uniporter
- PMID: 28325874
- PMCID: PMC5389301
- DOI: 10.1073/pnas.1620316114
Ion and inhibitor binding of the double-ring ion selectivity filter of the mitochondrial calcium uniporter
Abstract
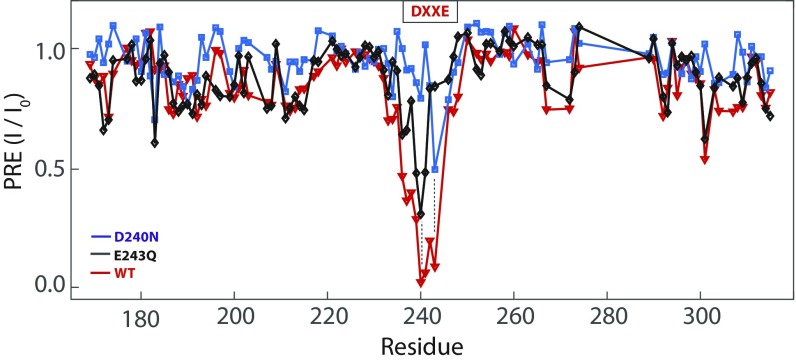
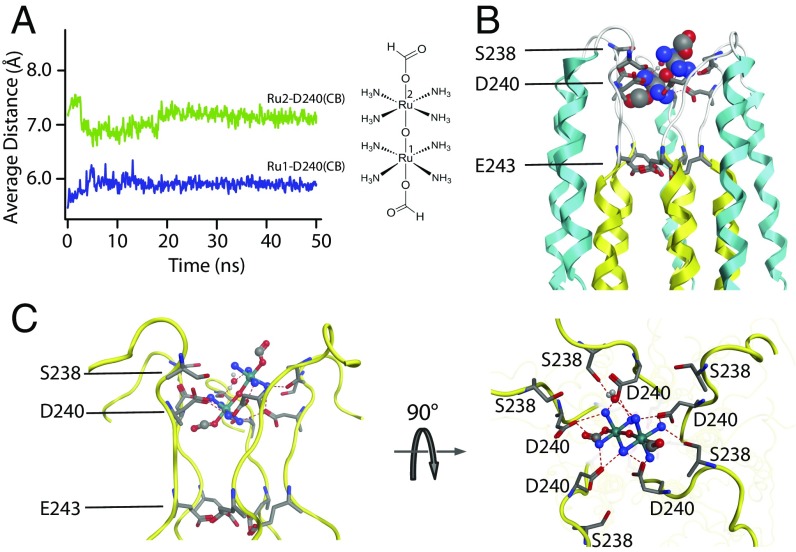
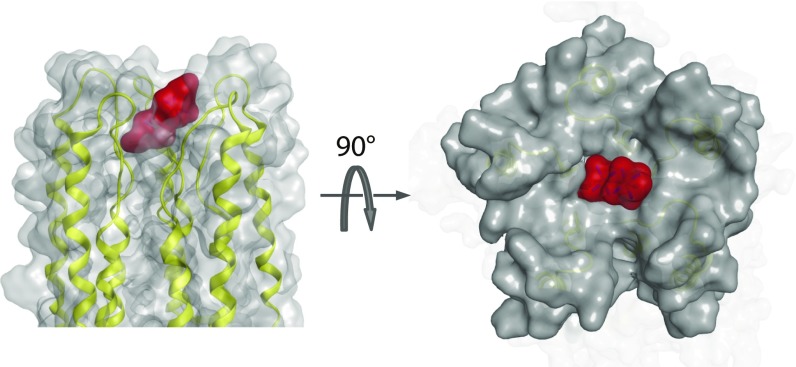
The calcium (Ca2+) uniporter of mitochondria is a holocomplex consisting of the Ca2+-conducting channel, known as mitochondrial calcium uniporter (MCU), and several accessory and regulatory components. A previous electrophysiology study found that the uniporter has high Ca2+ selectivity and conductance and this depends critically on the conserved amino acid sequence motif, DXXE (Asp-X-X-Glu) of MCU. A recent NMR structure of the MCU channel from Caenorhabditis elegans revealed that the DXXE forms two parallel carboxylate rings at the channel entrance that seem to serve as the ion selectivity filter, although direct ion interaction of this structural motif has not been addressed. Here, we use a paramagnetic probe, manganese (Mn2+), to investigate ion and inhibitor binding of this putative selectivity filter. Our paramagnetic NMR data show that mutants with a single carboxylate ring, NXXE (Asn-X-X-Glu) and DXXQ (Asp-X-X-Gln), each can bind Mn2+ specifically, whereas in the WT the two rings bind Mn2+ cooperatively, resulting in ∼1,000-fold higher apparent affinity. Ca2+ can specifically displace the bound Mn2+ at the DXXE site in the channel. Furthermore, titrating the sample with the known channel inhibitor ruthenium 360 (Ru360) can displace Mn2+ binding from the solvent-accessible Asp site but not the inner Glu site. The NMR titration data, together with structural analysis of the DXXE motif and molecular dynamics simulation, indicate that the double carboxylate rings at the apex of the MCU pore constitute the ion selectivity filter and that Ru360 directly blocks ion entry into the filter by binding to the outer carboxylate ring.
Keywords: MCU; NMR; Ru360 binding; calcium channel; selectivity filter.
Conflict of interest statement
The authors declare no conflict of interest.
Figures










References
-
- Vasington FD, Murphy JV. Ca ion uptake by rat kidney mitochondria and its dependence on respiration and phosphorylation. J Biol Chem. 1962;237(13925019):2670–2677. - PubMed
-
- Denton RM, McCormack JG. The role of calcium in the regulation of mitochondrial metabolism. Biochem Soc Trans. 1980;8(3):266–268. - PubMed
-
- Gunter TE, Pfeiffer DR. Mechanisms by which mitochondria transport calcium. Am J Physiol. 1990;258(5 Pt 1):C755–C786. - PubMed
-
- Kirichok Y, Krapivinsky G, Clapham DE. The mitochondrial calcium uniporter is a highly selective ion channel. Nature. 2004;427(6972):360–364. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
