

Acoustic startle modification as a tool for evaluating auditory function of the mouse: Progress, pitfalls, and potential
- PMID: 28327385
- PMCID: PMC5446932
- DOI: 10.1016/j.neubiorev.2017.03.009
Acoustic startle modification as a tool for evaluating auditory function of the mouse: Progress, pitfalls, and potential
Abstract
Acoustic startle response (ASR) modification procedures, especially prepulse inhibition (PPI), are increasingly used as behavioral measures of auditory processing and sensorimotor gating in rodents due to their perceived ease of implementation and short testing times. In practice, ASR and PPI procedures are extremely variable across animals, experimental setups, and studies, and the interpretation of results is subject to numerous caveats and confounding influences. We review considerations for modification of the ASR using acoustic stimuli, and we compare the sensitivity of PPI procedures to more traditional operant psychoacoustic techniques. We also discuss non-auditory variables that must be considered. We conclude that ASR and PPI measures cannot substitute for traditional operant techniques due to their low sensitivity. Additionally, a substantial amount of pilot testing must be performed to properly optimize an ASR modification experiment, negating any time benefit over operant conditioning. Nevertheless, there are some circumstances where ASR measures may be the only option for assessing auditory behavior, such as when testing mouse strains with early-onset hearing loss or learning impairments.
Copyright © 2017 Elsevier Ltd. All rights reserved.
Figures
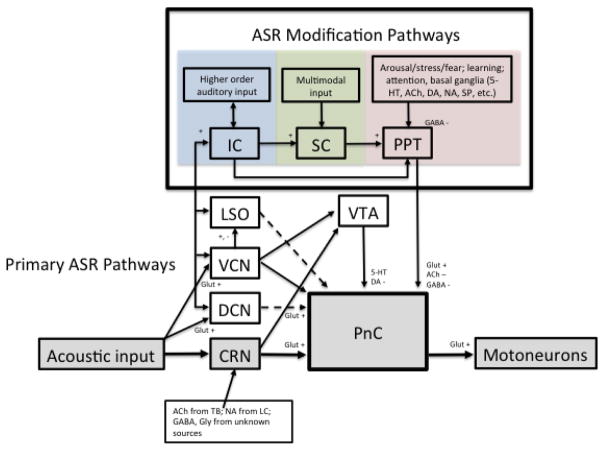
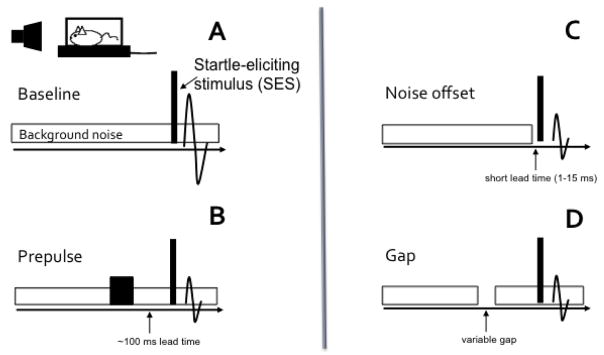
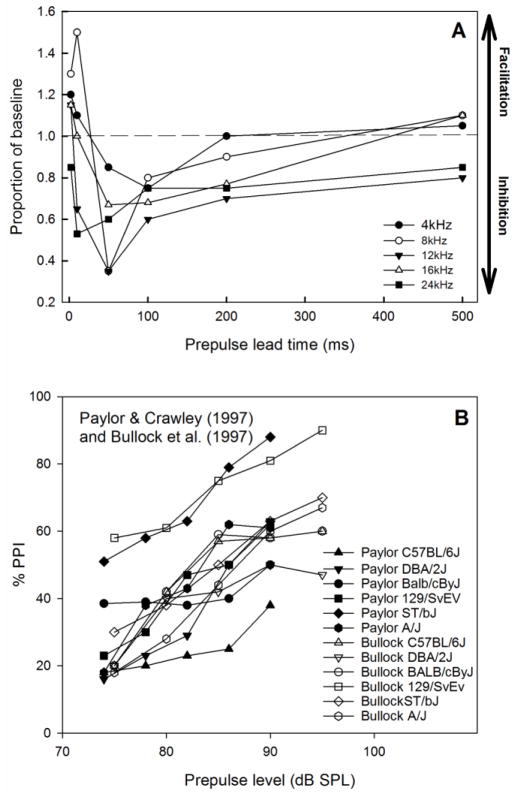
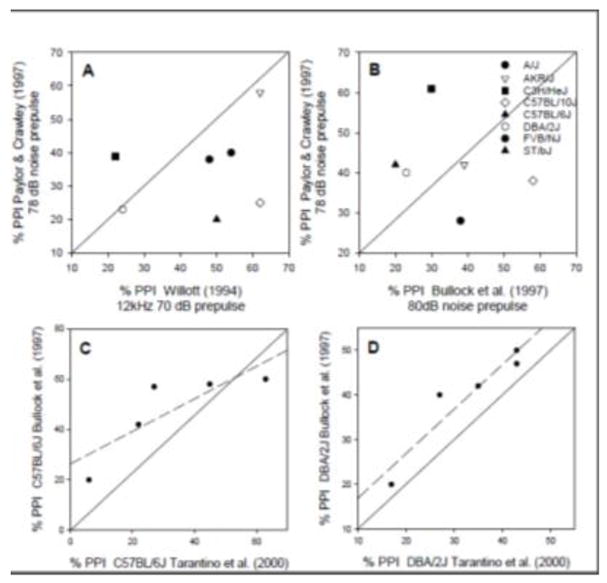
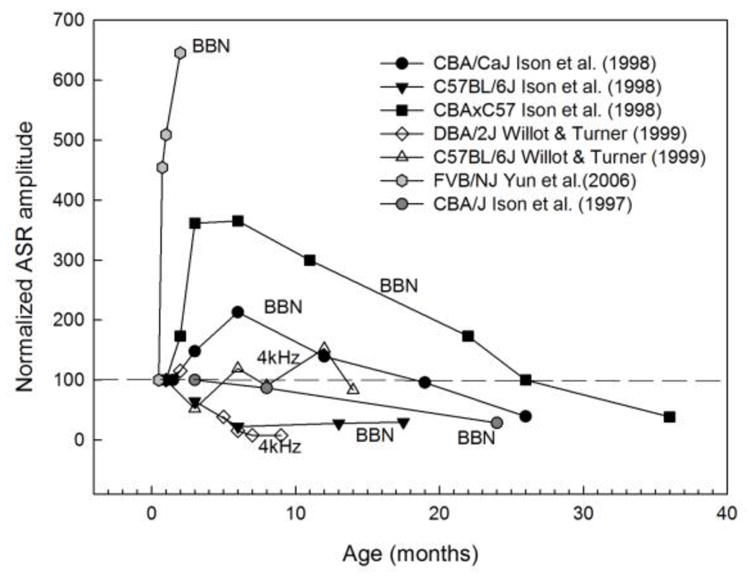
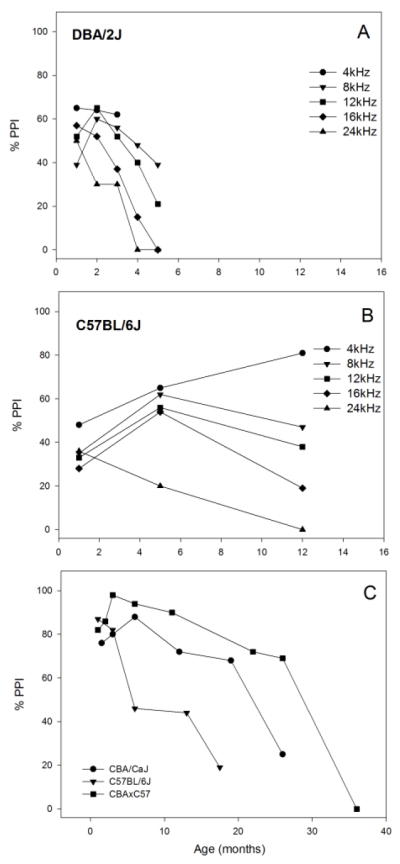
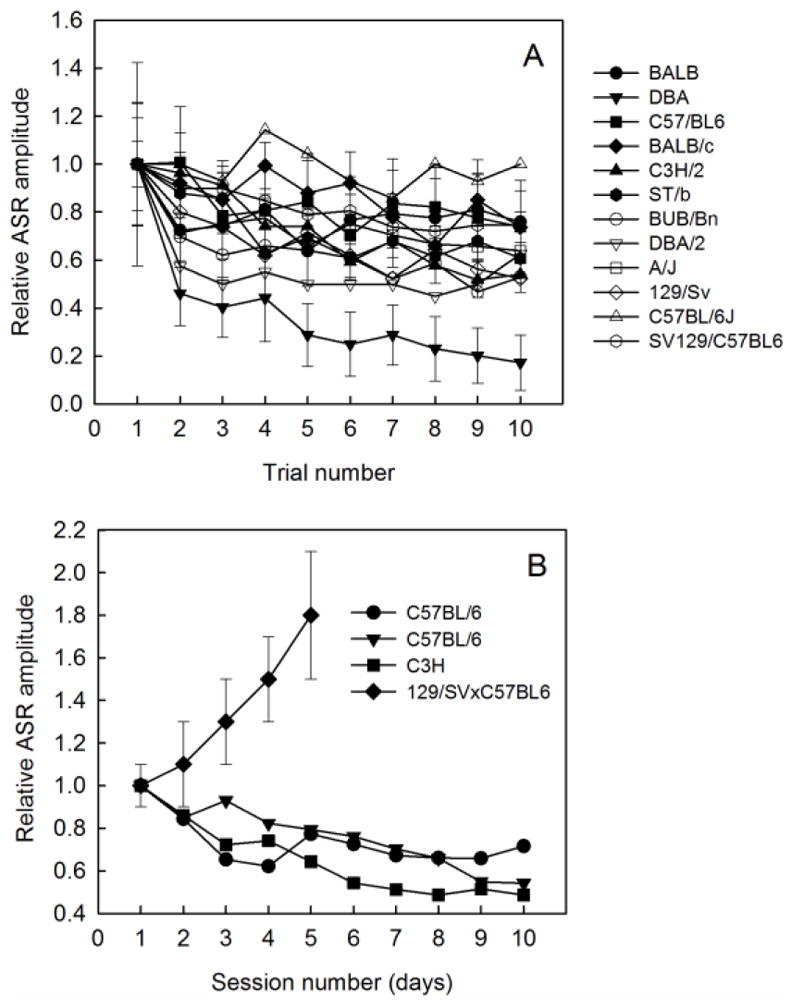
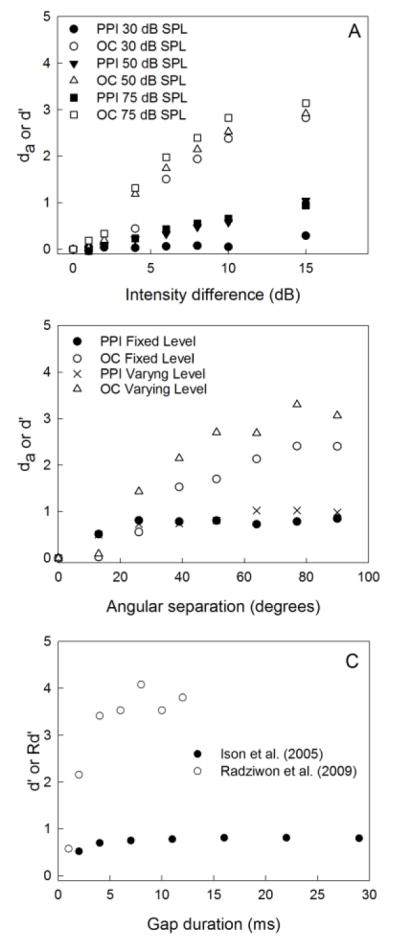
Similar articles
-
Relationships between the acoustic startle response and prepulse inhibition in C57BL/6J mice: a large-scale meta-analytic study.Mol Brain. 2018 Jul 13;11(1):42. doi: 10.1186/s13041-018-0382-7. Mol Brain. 2018. PMID: 30001725 Free PMC article.
-
Genetic reduction of MMP-9 in the Fmr1 KO mouse partially rescues prepulse inhibition of acoustic startle response.Brain Res. 2019 Sep 15;1719:24-29. doi: 10.1016/j.brainres.2019.05.029. Epub 2019 May 22. Brain Res. 2019. PMID: 31128097 Free PMC article.
-
Effects of age-related hearing loss on startle reflex and prepulse inhibition in mice on pure and mixed C57BL and 129 genetic background.Behav Brain Res. 2006 Sep 25;172(2):307-15. doi: 10.1016/j.bbr.2006.05.018. Epub 2006 Jun 30. Behav Brain Res. 2006. PMID: 16814879
-
Revisiting Psychoacoustic Methods for the Assessment of Fish Hearing.Adv Exp Med Biol. 2016;877:157-84. doi: 10.1007/978-3-319-21059-9_8. Adv Exp Med Biol. 2016. PMID: 26515314 Review.
-
Prepulse inhibition of the acoustic startle reflex and P50 gating in aging and alzheimer's disease.Ageing Res Rev. 2020 May;59:101028. doi: 10.1016/j.arr.2020.101028. Epub 2020 Feb 21. Ageing Res Rev. 2020. PMID: 32092463 Review.
Cited by
-
Making translation work: Harmonizing cross-species methodology in the behavioural neuroscience of Pavlovian fear conditioning.Neurosci Biobehav Rev. 2019 Dec;107:329-345. doi: 10.1016/j.neubiorev.2019.09.020. Epub 2019 Sep 12. Neurosci Biobehav Rev. 2019. PMID: 31521698 Free PMC article. Review.
-
The Shank3-InsG3680(+/+) mouse model of autism spectrum disorder displays auditory avoidance in a novel behavioral test.Front Behav Neurosci. 2023 Aug 24;17:1205507. doi: 10.3389/fnbeh.2023.1205507. eCollection 2023. Front Behav Neurosci. 2023. PMID: 37693284 Free PMC article.
-
Sex-Specific Role for Dopamine Receptor D2 in Dorsal Raphe Serotonergic Neuron Modulation of Defensive Acoustic Startle and Dominance Behavior.eNeuro. 2020 Nov 19;7(6):ENEURO.0202-20.2020. doi: 10.1523/ENEURO.0202-20.2020. Print 2020 Nov-Dec. eNeuro. 2020. PMID: 33214315 Free PMC article.
-
A genetically identified population of layer 4 neurons in auditory cortex that contributes to pre-pulse inhibition of the acoustic startle response.Front Neural Circuits. 2022 Sep 8;16:972157. doi: 10.3389/fncir.2022.972157. eCollection 2022. Front Neural Circuits. 2022. PMID: 36160948 Free PMC article.
-
Sound-seeking before and after hearing loss in mice.Sci Rep. 2024 Aug 19;14(1):19181. doi: 10.1038/s41598-024-67577-7. Sci Rep. 2024. PMID: 39160202 Free PMC article.
References
-
- Adams AL, Hudson A, Ryan CL, Doucette TA. Effects of estrous stage and time of day on prepulse inhibition in female rats. Journal of neuroscience methods. 2008 Aug 30;173(2):295–8. - PubMed
-
- Altschuler RA, Dolan DF, Halsey K, Kanicki A, Deng N, Martin C, Eberle J, Kohrman DC, Miller RA, Schacht J. Age-related changes in auditory nerve–inner hair cell connections, hair cell numbers, auditory brain stem response and gap detection in UM-HET4 mice. Neuroscience. 2015 Apr 30;292:22–33. - PMC - PubMed
-
- Amanipour R, Cresoe S, Borlongan C, Frisina R, Walton J. Effects of Mild Traumatic Brain Injury on Auditory Function in a Mouse Model. Biomedical Engineering Conference (SBEC), 2016 32nd Southern; 2016 Mar 11; IEEE; pp. 13–14. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
