

Lineage-Biased Stem Cells Maintain Estrogen-Receptor-Positive and -Negative Mouse Mammary Luminal Lineages
- PMID: 28329676
- PMCID: PMC5408864
- DOI: 10.1016/j.celrep.2017.02.071
Lineage-Biased Stem Cells Maintain Estrogen-Receptor-Positive and -Negative Mouse Mammary Luminal Lineages
Abstract
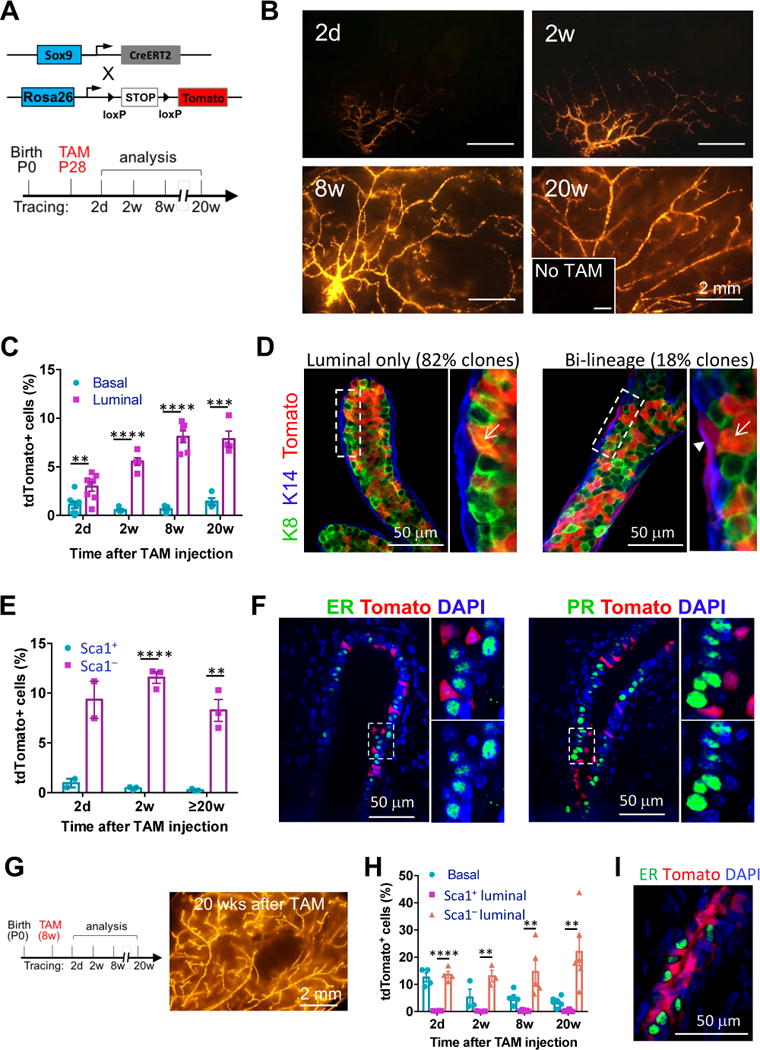
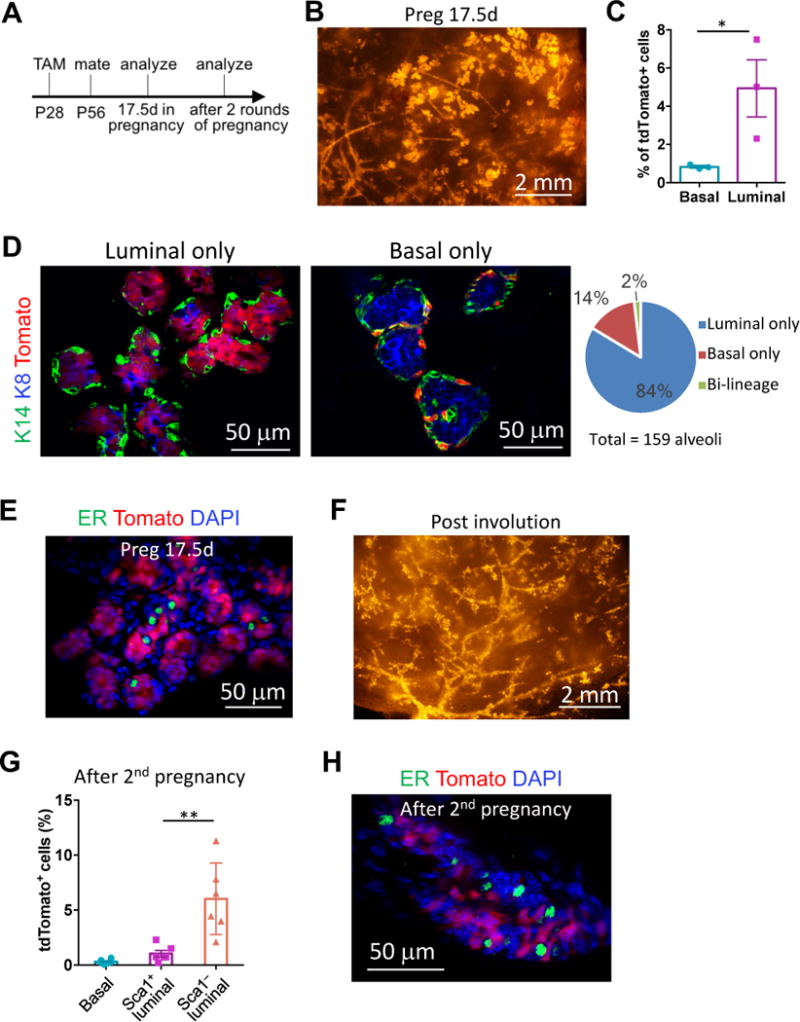
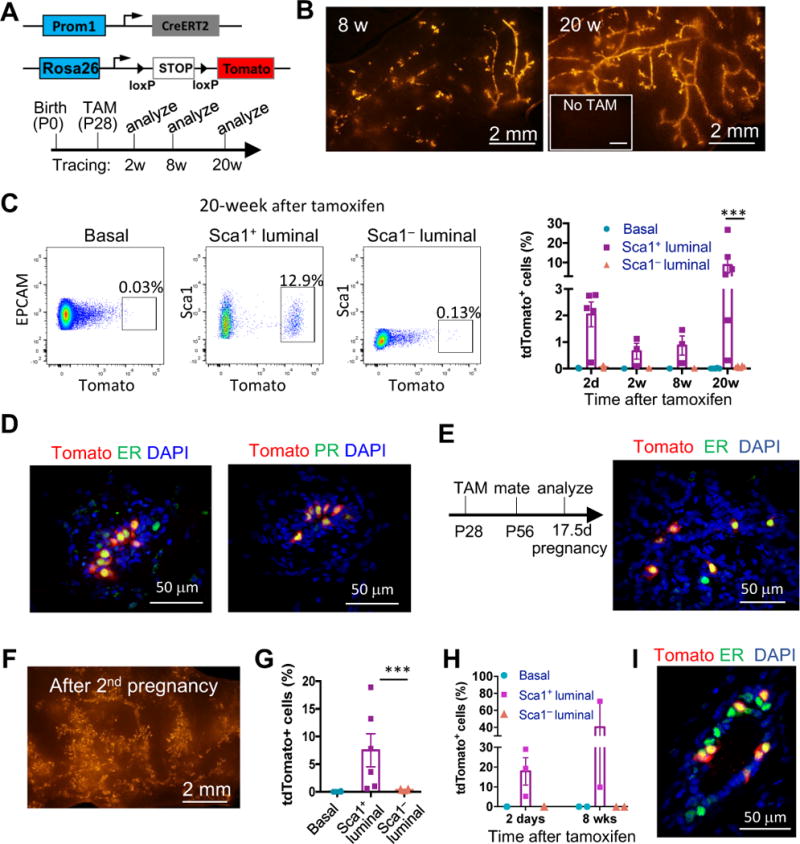
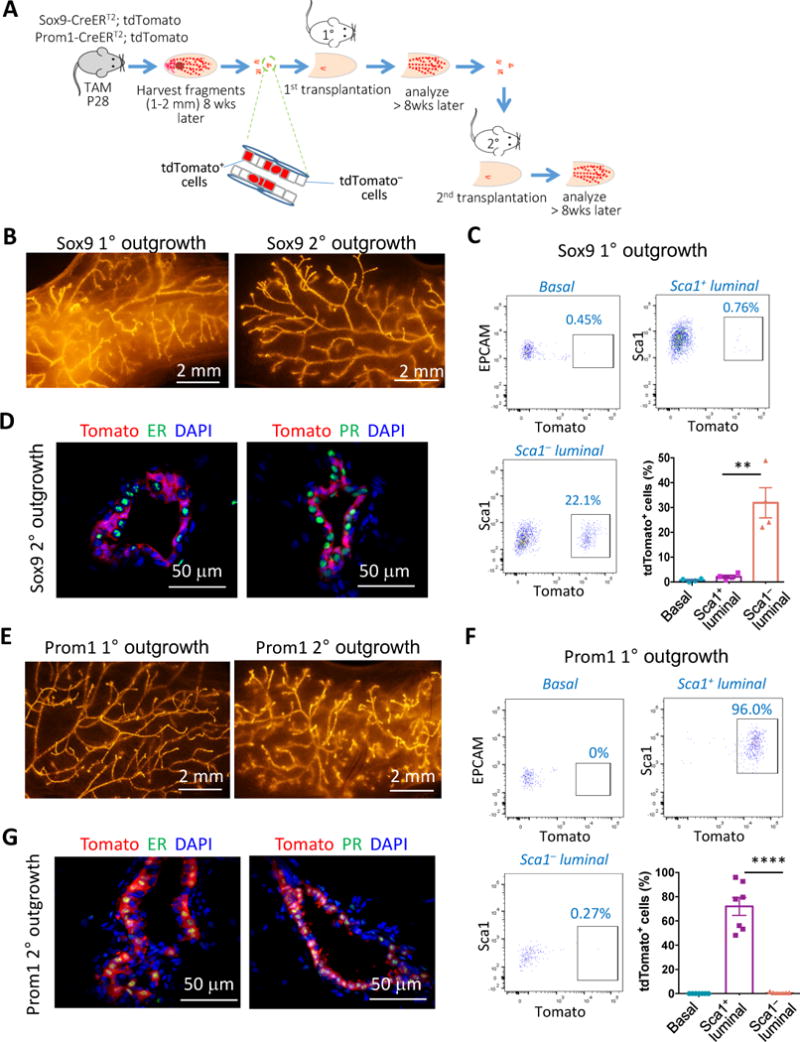
Delineating the mammary differentiation hierarchy is important for the study of mammary gland development and tumorigenesis. Mammary luminal cells are considered a major origin of human breast cancers. However, how estrogen-receptor-positive (ER+) and ER- luminal cells are developed and maintained remains poorly understood. The prevailing model suggests that a common stem/progenitor cell generates both cell types. Through genetic lineage tracing in mice, we find that SOX9-expressing cells specifically contribute to the development and maintenance of ER- luminal cells and, to a lesser degree, basal cells. In parallel, PROM1-expressing cells give rise only to ER+ luminal cells. Both SOX9+ and PROM1+ cells specifically sustain their respective lineages even after pregnancy-caused tissue remodeling or serial transplantation, demonstrating characteristic properties of long-term repopulating stem cells. Thus, our data reveal that mouse mammary ER+ and ER- luminal cells are two independent lineages that are maintained by distinct stem cells, providing a revised mammary epithelial cell hierarchy.
Keywords: breast cancer; cancer cell-of-origin; estrogen receptor negative cancer; estrogen receptor positive cancer; lineage tracing; mammary differentiation; mammary gland development; mammary gland regeneration; mammary stem cells; stem cell hierarchy.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- de Visser KE, Ciampricotti M, Michalak EM, Tan DW, Speksnijder EN, Hau CS, Clevers H, Barker N, Jonkers J. Developmental stage-specific contribution of LGR5(+) cells to basal and luminal epithelial lineages in the postnatal mammary gland. J Pathol. 2012;228:300–309. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
