

Efficient Ex Vivo Engineering and Expansion of Highly Purified Human Hematopoietic Stem and Progenitor Cell Populations for Gene Therapy
- PMID: 28330619
- PMCID: PMC5390102
- DOI: 10.1016/j.stemcr.2017.02.010
Efficient Ex Vivo Engineering and Expansion of Highly Purified Human Hematopoietic Stem and Progenitor Cell Populations for Gene Therapy
Abstract
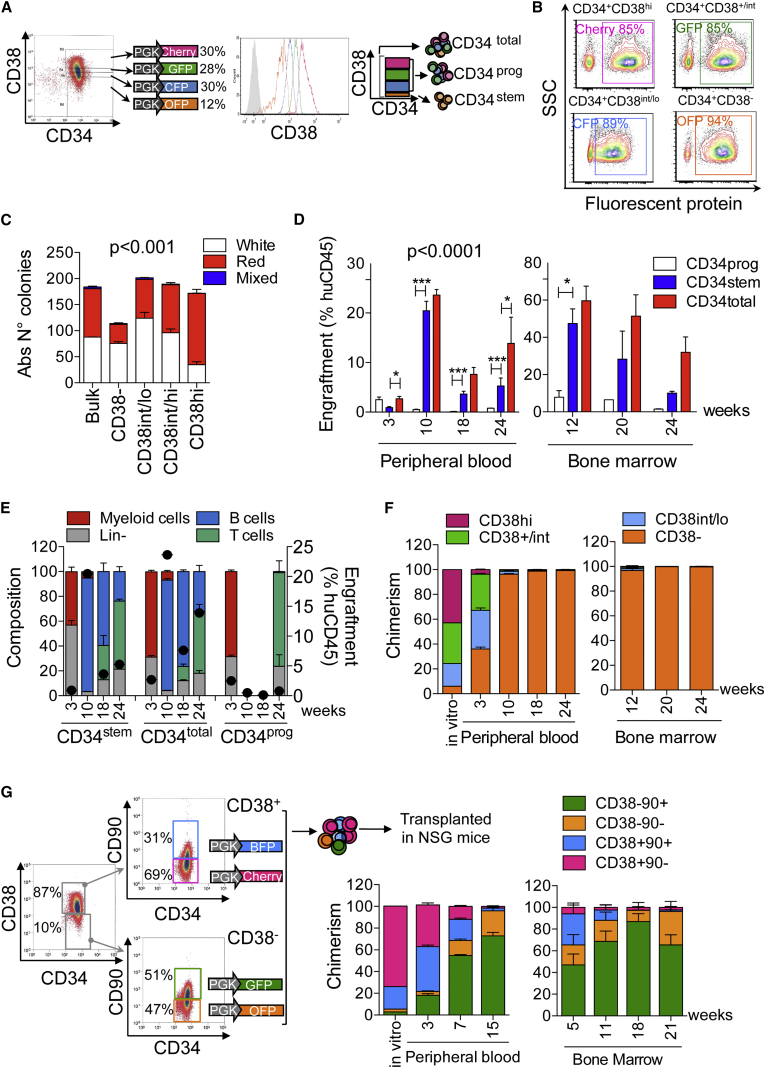
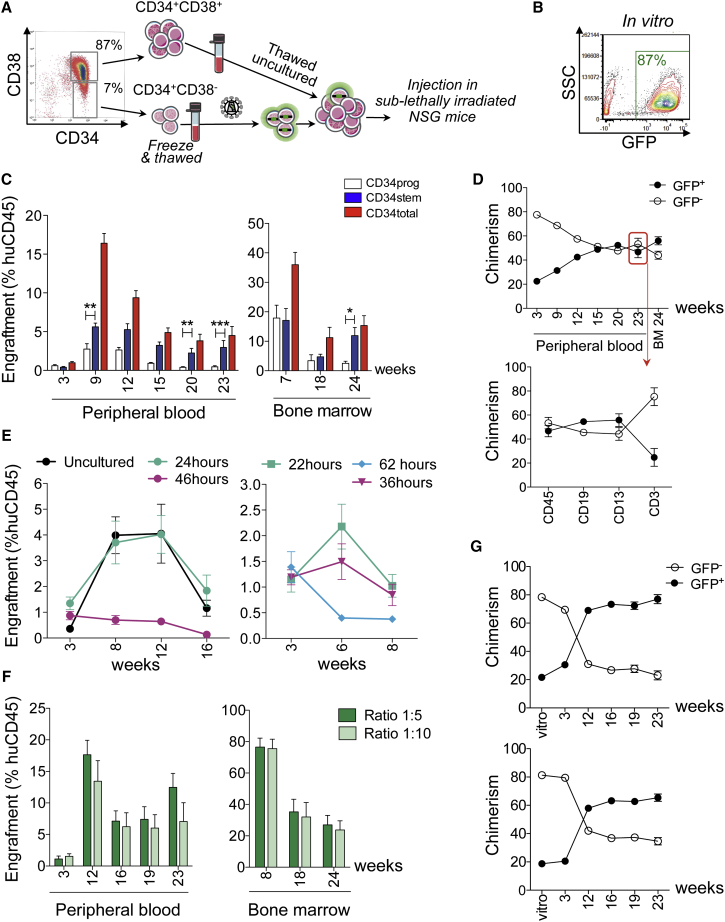
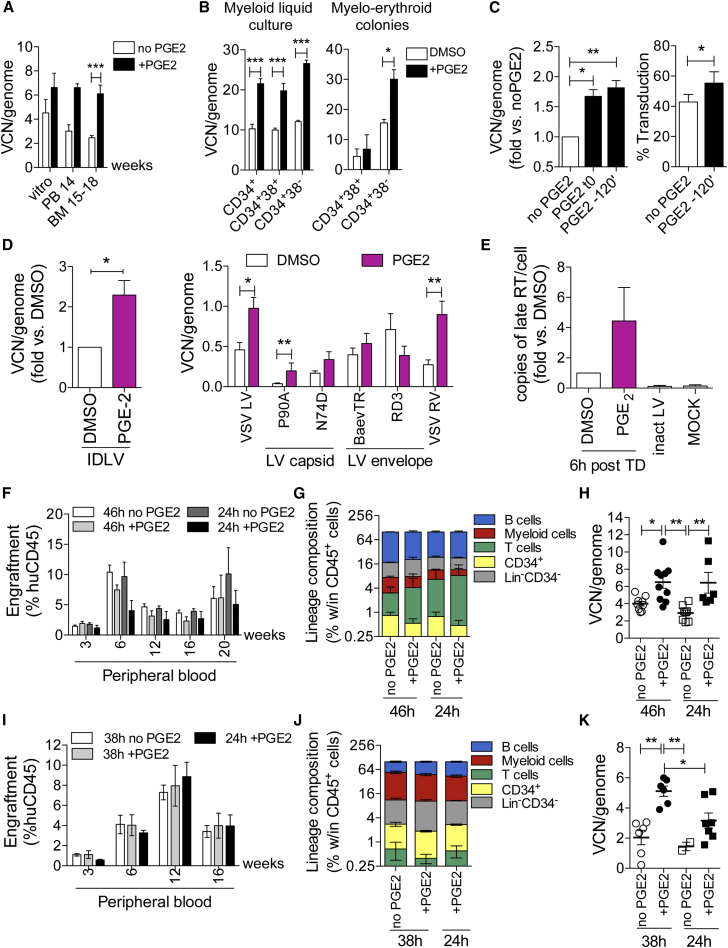
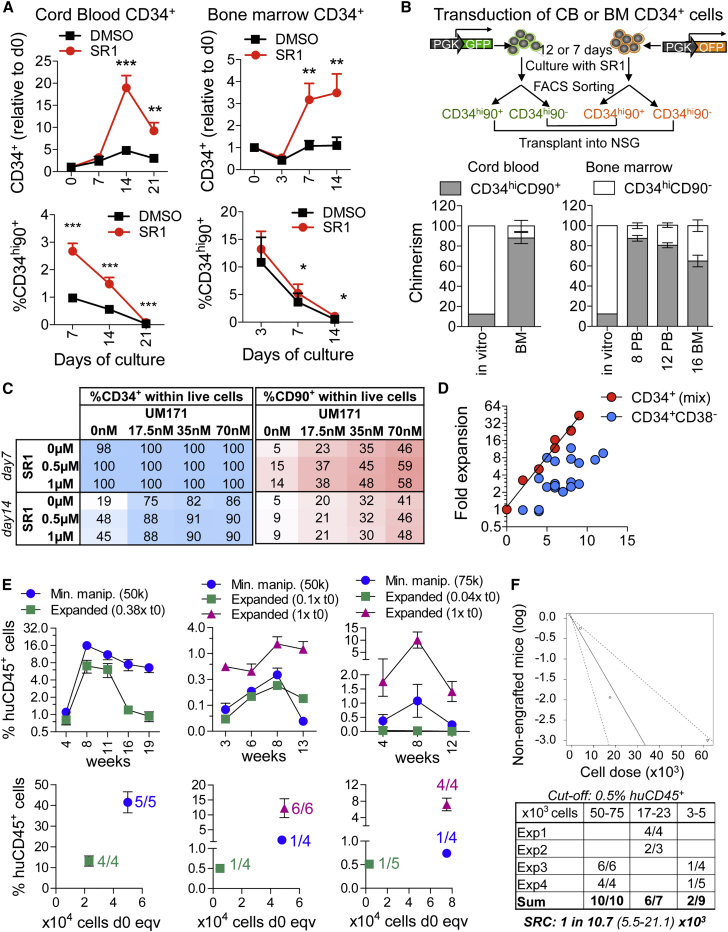
Ex vivo gene therapy based on CD34+ hematopoietic stem cells (HSCs) has shown promising results in clinical trials, but genetic engineering to high levels and in large scale remains challenging. We devised a sorting strategy that captures more than 90% of HSC activity in less than 10% of mobilized peripheral blood (mPB) CD34+ cells, and modeled a transplantation protocol based on highly purified, genetically engineered HSCs co-infused with uncultured progenitor cells. Prostaglandin E2 stimulation allowed near-complete transduction of HSCs with lentiviral vectors during a culture time of less than 38 hr, mitigating the negative impact of standard culture on progenitor cell function. Exploiting the pyrimidoindole derivative UM171, we show that transduced mPB CD34+CD38- cells with repopulating potential could be expanded ex vivo. Implementing these findings in clinical gene therapy protocols will improve the efficacy, safety, and sustainability of gene therapy and generate new opportunities in the field of gene editing.
Keywords: HSC expansion; HSC gene therapy; UM171; lentiviral vector transduction; prostaglandin E2; purified HSCs.
Copyright © 2017 The Author(s). Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Biasco L., Pellin D., Scala S., Dionisio F., Basso-Ricci L., Leonardelli L., Scaramuzza S., Baricordi C., Ferrua F., Cicalese M.P. In vivo tracking of human hematopoiesis reveals patterns of clonal dynamics during early and steady-state reconstitution phases. Cell Stem Cell. 2015;19:107–119. - PMC - PubMed
-
- Biffi A., Montini E., Lorioli L., Cesani M., Fumagalli F., Plati T., Baldoli C., Martino S., Calabria A., Canale S. Lentiviral hematopoietic stem cell gene therapy benefits metachromatic leukodystrophy. Science. 2013;341:1233158. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
