

Cell-Specific Variation in E-Selectin Ligand Expression among Human Peripheral Blood Mononuclear Cells: Implications for Immunosurveillance and Pathobiology
- PMID: 28330896
- PMCID: PMC5426364
- DOI: 10.4049/jimmunol.1601636
Cell-Specific Variation in E-Selectin Ligand Expression among Human Peripheral Blood Mononuclear Cells: Implications for Immunosurveillance and Pathobiology
Abstract
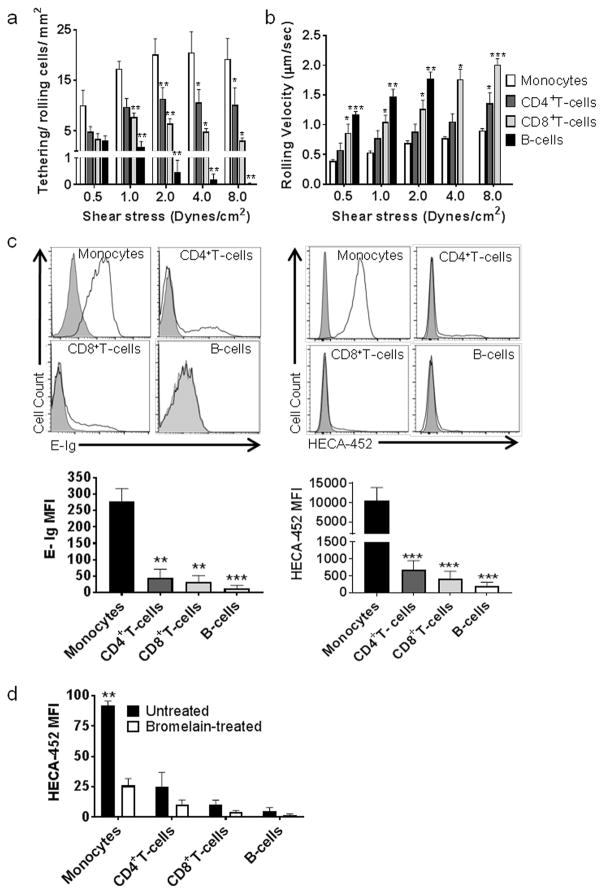
Both host defense and immunopathology are shaped by the ordered recruitment of circulating leukocytes to affected sites, a process initiated by binding of blood-borne cells to E-selectin displayed at target endothelial beds. Accordingly, knowledge of the expression and function of leukocyte E-selectin ligands is key to understanding the tempo and specificity of immunoreactivity. In this study, we performed E-selectin adherence assays under hemodynamic flow conditions coupled with flow cytometry and Western blot analysis to elucidate the function and structural biology of glycoprotein E-selectin ligands expressed on human PBMCs. Circulating monocytes uniformly express high levels of the canonical E-selectin binding determinant sialyl Lewis X (sLeX) and display markedly greater adhesive interactions with E-selectin than do circulating lymphocytes, which exhibit variable E-selectin binding among CD4+ and CD8+ T cells but no binding by B cells. Monocytes prominently present sLeX decorations on an array of protein scaffolds, including P-selectin glycoprotein ligand-1, CD43, and CD44 (rendering the E-selectin ligands cutaneous lymphocyte Ag, CD43E, and hematopoietic cell E-selectin/L-selectin ligand, respectively), and B cells altogether lack E-selectin ligands. Quantitative PCR gene expression studies of glycosyltransferases that regulate display of sLeX reveal high transcript levels among circulating monocytes and low levels among circulating B cells, and, commensurately, cell surface α(1,3)-fucosylation reveals that acceptor sialyllactosaminyl glycans convertible into sLeX are abundantly expressed on human monocytes yet are relatively deficient on B cells. Collectively, these findings unveil distinct cell-specific patterns of E-selectin ligand expression among human PBMCs, indicating that circulating monocytes are specialized to engage E-selectin and providing key insights into the molecular effectors mediating recruitment of these cells at inflammatory sites.
Copyright © 2017 by The American Association of Immunologists, Inc.
Figures





References
-
- Angiari S. Selectin-mediated leukocyte trafficking during the development of autoimmune disease. Autoimmun Rev. 2015;14:984–995. - PubMed
-
- Ley K, Laudanna C, Cybulsky MI, Nourshargh S. Getting to the site of inflammation: the leukocyte adhesion cascade updated. Nature reviews Immunology. 2007;7:678–689. - PubMed
-
- Bevilacqua M, Butcher E, Furie B, Gallatin M, Gimbrone M, Harlan J, Kishimoto K, Lasky L, McEver R. Selectins: a family of adhesion receptors. Cell. 1991;67:233. - PubMed
-
- Yao L, Setiadi H, Xia L, Laszik Z, Taylor FB, McEver RP. Divergent inducible expression of P-selectin and E-selectin in mice and primates. Blood. 1999;94:3820–3828. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
