

Heterologous Packaging Signals on Segment 4, but Not Segment 6 or Segment 8, Limit Influenza A Virus Reassortment
- PMID: 28331085
- PMCID: PMC5432880
- DOI: 10.1128/JVI.00195-17
Heterologous Packaging Signals on Segment 4, but Not Segment 6 or Segment 8, Limit Influenza A Virus Reassortment
Abstract
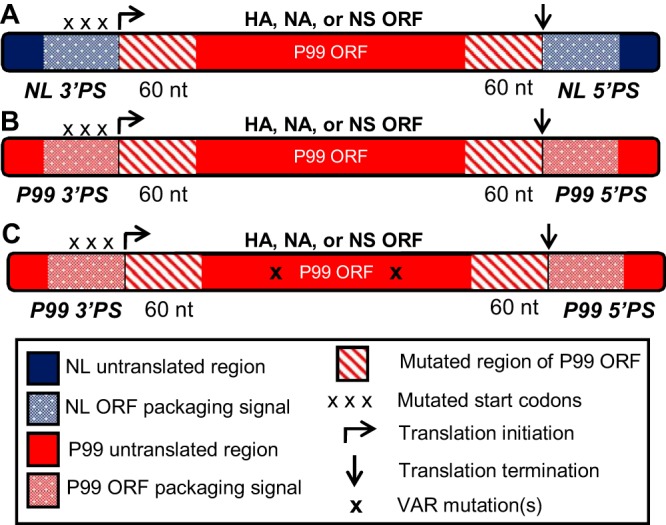
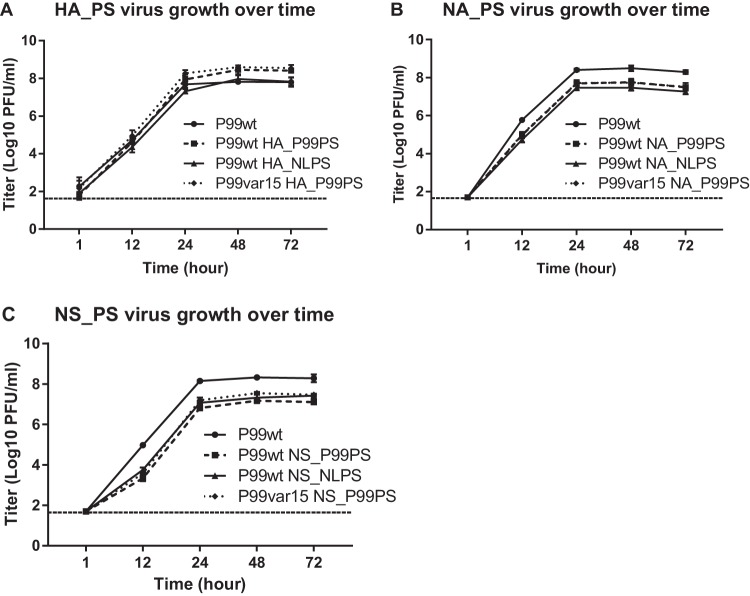
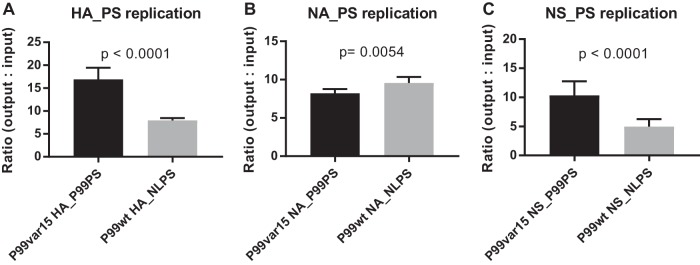
Influenza A virus (IAV) RNA packaging signals serve to direct the incorporation of IAV gene segments into virus particles, and this process is thought to be mediated by segment-segment interactions. These packaging signals are segment and strain specific, and as such, they have the potential to impact reassortment outcomes between different IAV strains. Our study aimed to quantify the impact of packaging signal mismatch on IAV reassortment using the human seasonal influenza A/Panama/2007/99 (H3N2) and pandemic influenza A/Netherlands/602/2009 (H1N1) viruses. Focusing on the three most divergent segments, we constructed pairs of viruses that encoded identical proteins but differed in the packaging signal regions on a single segment. We then evaluated the frequency with which segments carrying homologous versus heterologous packaging signals were incorporated into reassortant progeny viruses. We found that, when segment 4 (HA) of coinfecting parental viruses was modified, there was a significant preference for the segment containing matched packaging signals relative to the background of the virus. This preference was apparent even when the homologous HA constituted a minority of the HA segment population available in the cell for packaging. Conversely, when segment 6 (NA) or segment 8 (NS) carried modified packaging signals, there was no significant preference for homologous packaging signals. These data suggest that movement of NA and NS segments between the human H3N2 and H1N1 lineages is unlikely to be restricted by packaging signal mismatch, while movement of the HA segment would be more constrained. Our results indicate that the importance of packaging signals in IAV reassortment is segment dependent.IMPORTANCE Influenza A viruses (IAVs) can exchange genes through reassortment. This process contributes to both the highly diverse population of IAVs found in nature and the formation of novel epidemic and pandemic IAV strains. Our study sought to determine the extent to which IAV packaging signal divergence impacts reassortment between seasonal IAVs. Our knowledge in this area is lacking, and insight into the factors that influence IAV reassortment will inform and strengthen ongoing public health efforts to anticipate the emergence of new viruses. We found that the packaging signals on the HA segment, but not the NA or NS segments, restricted IAV reassortment. Thus, the packaging signals of the HA segment could be an important factor in determining the likelihood that two IAV strains of public health interest will undergo reassortment.
Keywords: evolution; influenza virus; packaging; reassortment; segment mismatch.
Copyright © 2017 American Society for Microbiology.
Figures

References
-
- Shaw ML, Palese P. 2013. Orthomyxoviridae, p 1151–1185. In Knipe DM, Howley PM, Cohen JI, Griffin DE, Lamb RA, Martin MA, Racaniello VR, Roizman B (ed), Fields virology, 6th ed, vol 1 Lippincott Williams & Wilkins, Philadelphia, PA.
-
- Garten RJ, Davis CT, Russell CA, Shu B, Lindstrom S, Balish A, Sessions WM, Xu X, Skepner E, Deyde V, Okomo-Adhiambo M, Gubareva L, Barnes J, Smith CB, Emery SL, Hillman MJ, Rivailler P, Smagala J, de Graaf M, Burke DF, Fouchier RAM, Pappas C, Alpuche-Aranda CM, López-Gatell H, Olivera H, López I, Myers CA, Faix D, Blair PJ, Yu C, Keene KM, Dotson PD, Boxrud D, Sambol AR, Abid SH, St. George K, Bannerman T, Moore AL, Stringer DJ, Blevins P, Demmler-Harrison GJ, Ginsberg M, Kriner P, Waterman S, Smole S, Guevara HF, Belongia EA, Clark PA, Beatrice ST, Donis R, Katz J, Finelli L, Bridges CB, Shaw M, Jernigan DB, Uyeki TM, Smith DJ, Klimov AI, Cox NJ. 2009. Antigenic and genetic characteristics of swine-origin 2009 A(H1N1) influenza viruses circulating in humans. Science 325:197–201. doi: 10.1126/science.1176225. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
