

Diversity, Distribution, and Evolution of Tomato Viruses in China Uncovered by Small RNA Sequencing
- PMID: 28331089
- PMCID: PMC5432854
- DOI: 10.1128/JVI.00173-17
Diversity, Distribution, and Evolution of Tomato Viruses in China Uncovered by Small RNA Sequencing
Abstract
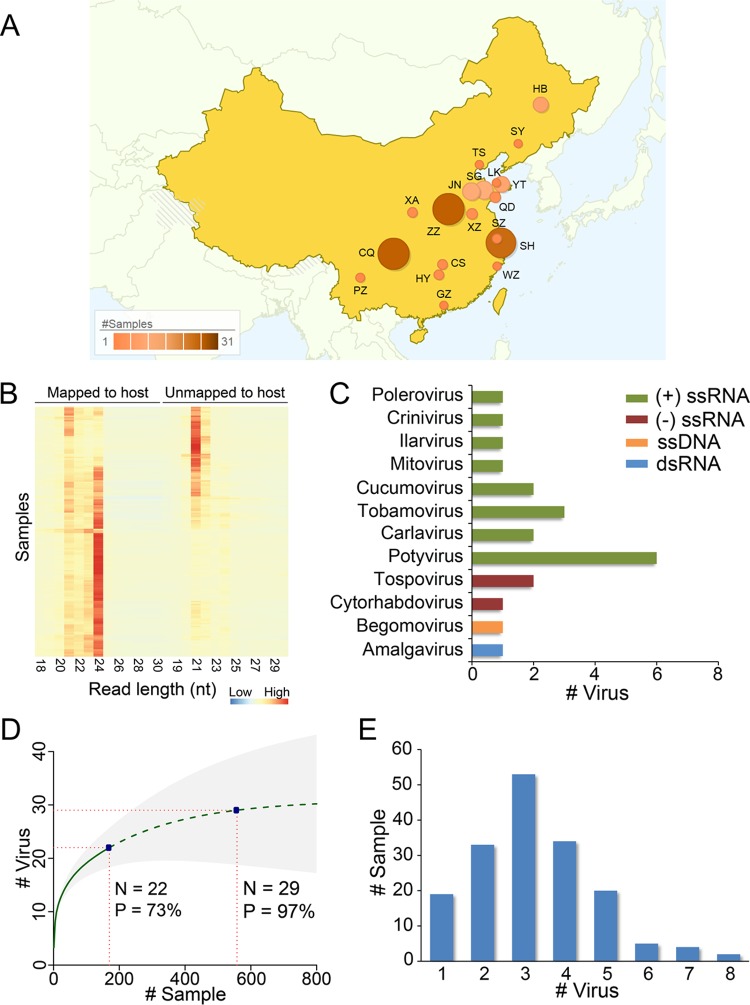
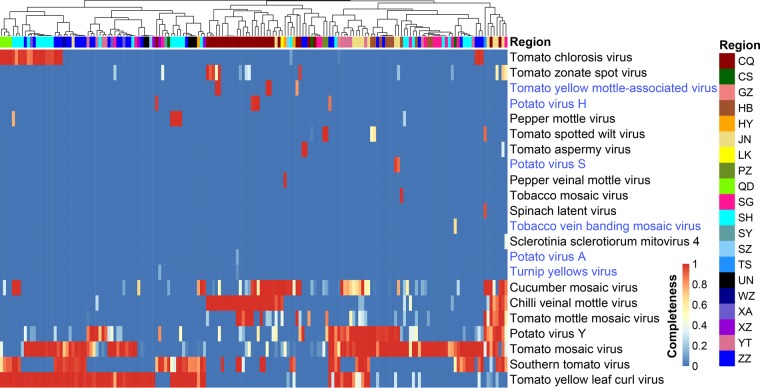
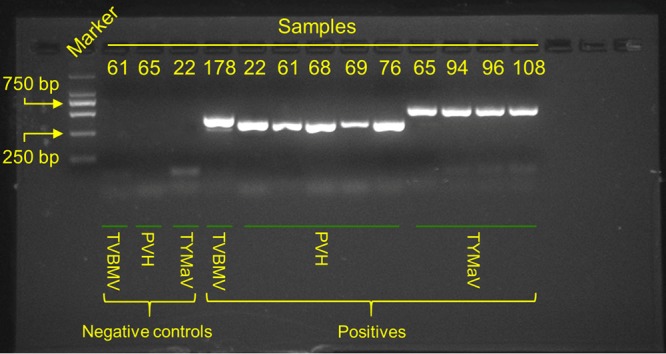
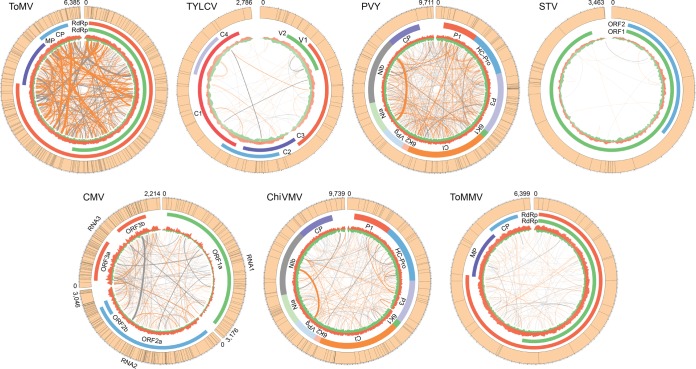
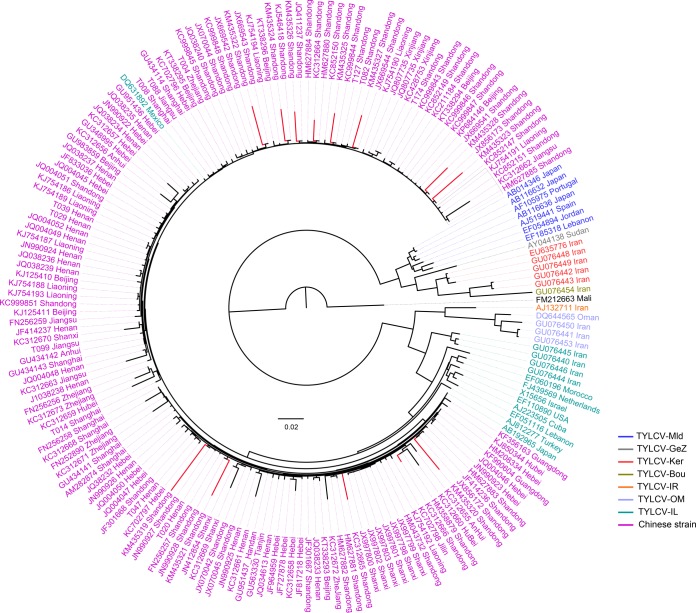
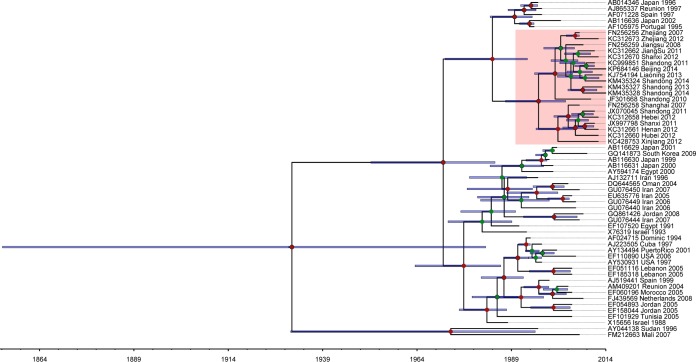
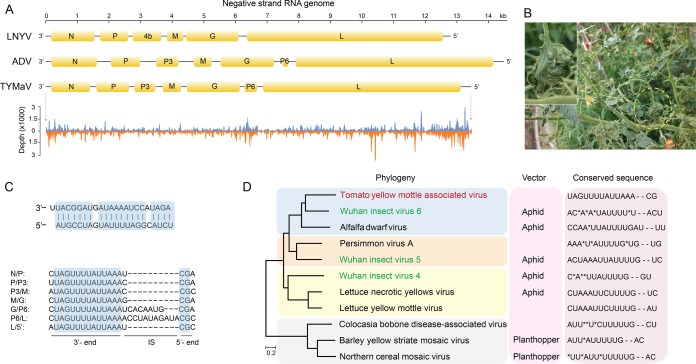
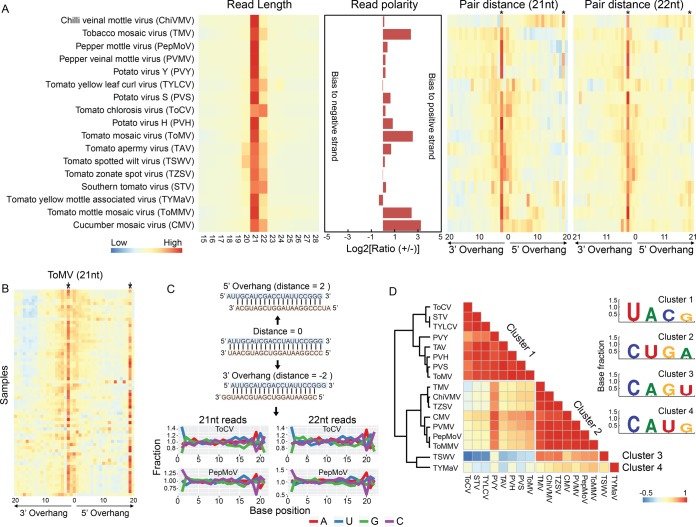
Tomato is a major vegetable crop that has tremendous popularity. However, viral disease is still a major factor limiting tomato production. Here, we report the tomato virome identified through sequencing small RNAs of 170 field-grown samples collected in China. A total of 22 viruses were identified, including both well-documented and newly detected viruses. The tomato viral community is dominated by a few species, and they exhibit polymorphisms and recombination in the genomes with cold spots and hot spots. Most samples were coinfected by multiple viruses, and the majority of identified viruses are positive-sense single-stranded RNA viruses. Evolutionary analysis of one of the most dominant tomato viruses, Tomato yellow leaf curl virus (TYLCV), predicts its origin and the time back to its most recent common ancestor. The broadly sampled data have enabled us to identify several unreported viruses in tomato, including a completely new virus, which has a genome of ∼13.4 kb and groups with aphid-transmitted viruses in the genus Cytorhabdovirus Although both DNA and RNA viruses can trigger the biogenesis of virus-derived small interfering RNAs (vsiRNAs), we show that features such as length distribution, paired distance, and base selection bias of vsiRNA sequences reflect different plant Dicer-like proteins and Argonautes involved in vsiRNA biogenesis. Collectively, this study offers insights into host-virus interaction in tomato and provides valuable information to facilitate the management of viral diseases.IMPORTANCE Tomato is an important source of micronutrients in the human diet and is extensively consumed around the world. Virus is among the major constraints on tomato production. Categorizing virus species that are capable of infecting tomato and understanding their diversity and evolution are challenging due to difficulties in detecting such fast-evolving biological entities. Here, we report the landscape of the tomato virome in China, the leading country in tomato production. We identified dozens of viruses present in tomato, including both well-documented and completely new viruses. Some newly emerged viruses in tomato were found to spread fast, and therefore, prompt attention is needed to control them. Moreover, we show that the virus genomes exhibit considerable degree of polymorphisms and recombination, and the virus-derived small interfering RNA (vsiRNA) sequences indicate distinct vsiRNA biogenesis mechanisms for different viruses. The Chinese tomato virome that we developed provides valuable information to facilitate the management of tomato viral diseases.
Keywords: small RNA sequencing; tomato virome; viral diversity and evolution.
Copyright © 2017 American Society for Microbiology.
Figures
References
-
- Bani R, Josiah M, Kra E. 2006. Postharvest losses of tomatoes in transit. Agric Mech Asia Afr Lat Am 37:84–86.
-
- Brunt A, Crabtree K, Dallwitz M, Gibbs A, Watson L, Zurcher E (ed). 1997. Plant viruses online: descriptions and lists from the VIDE database. Version 16 January 1997 Australian National University, Canberra, Australia: http://sdb.im.ac.cn/vide/refs.htm.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
