

The epigenetic landscape related to reactive oxygen species formation in the cardiovascular system
- PMID: 28332701
- PMCID: PMC5446579
- DOI: 10.1111/bph.13792
The epigenetic landscape related to reactive oxygen species formation in the cardiovascular system
Abstract
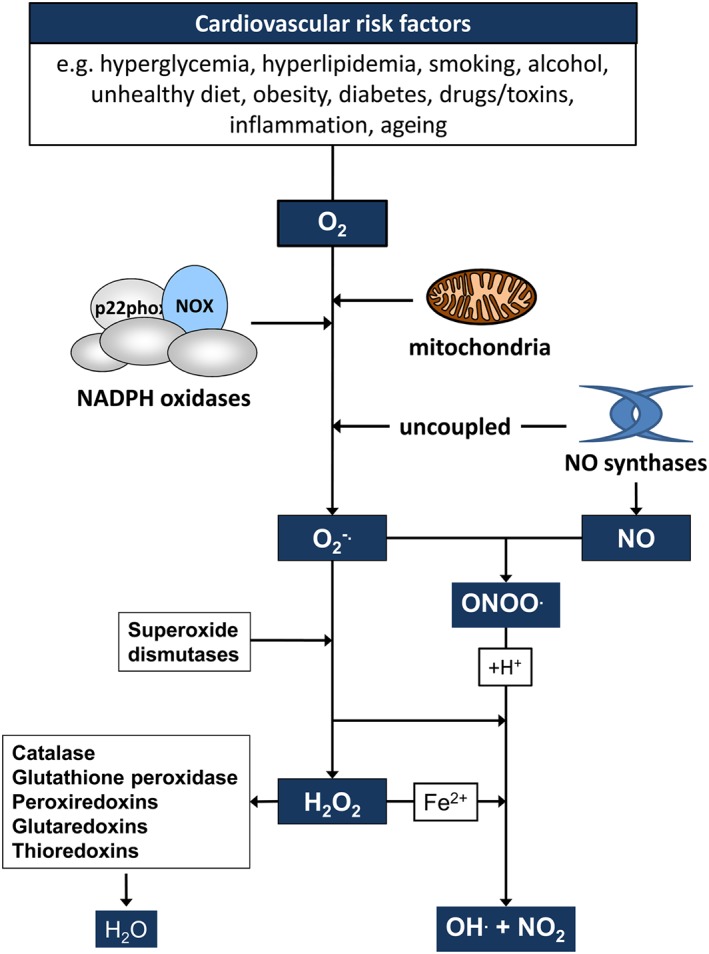
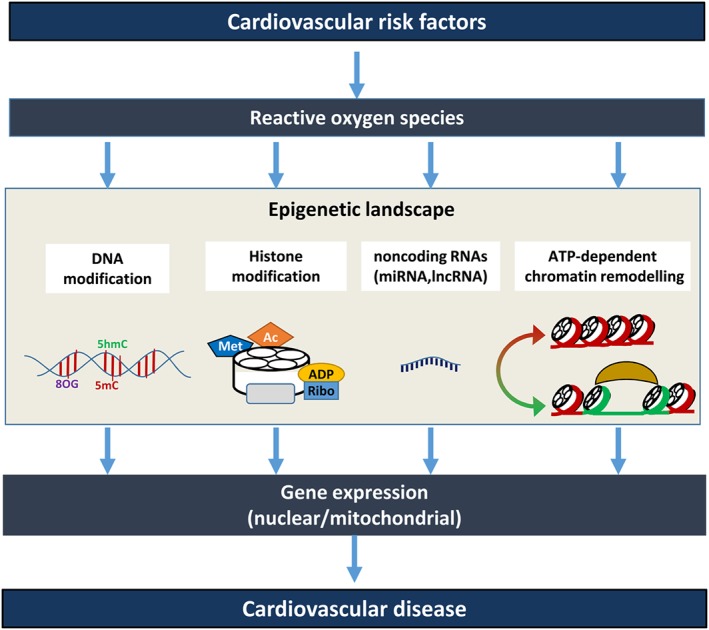
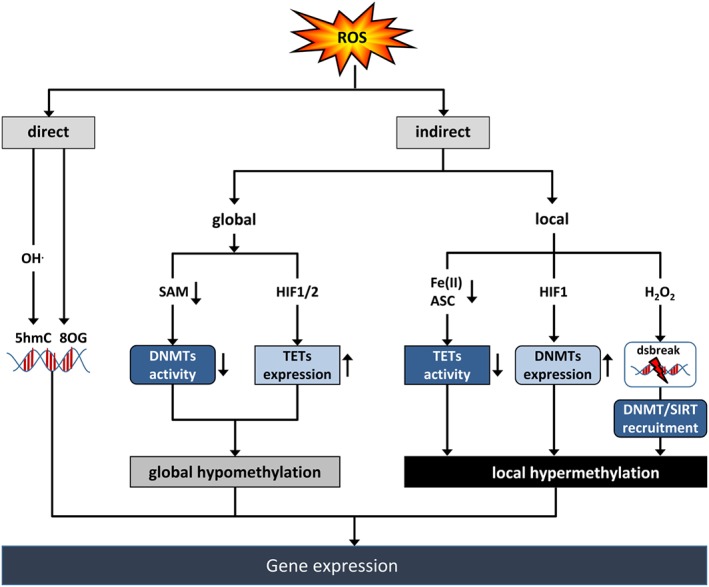
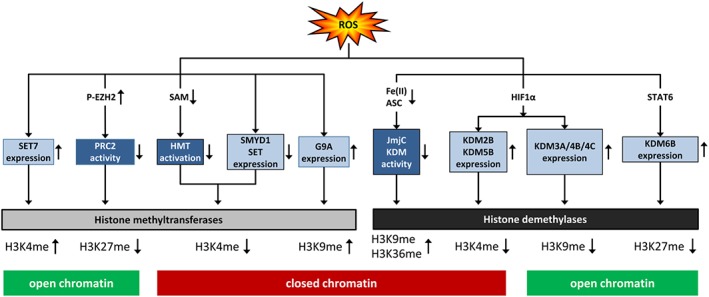
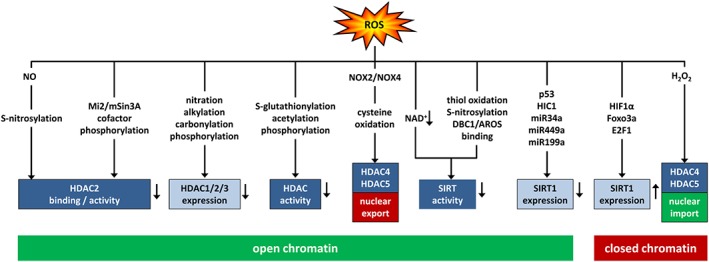
Cardiovascular diseases are among the leading causes of death worldwide. Reactive oxygen species (ROS) can act as damaging molecules but also represent central hubs in cellular signalling networks. Increasing evidence indicates that ROS play an important role in the pathogenesis of cardiovascular diseases, although the underlying mechanisms and consequences of pathophysiologically elevated ROS in the cardiovascular system are still not completely resolved. More recently, alterations of the epigenetic landscape, which can affect DNA methylation, post-translational histone modifications, ATP-dependent alterations to chromatin and non-coding RNA transcripts, have been considered to be of increasing importance in the pathogenesis of cardiovascular diseases. While it has long been accepted that epigenetic changes are imprinted during development or even inherited and are not changed after reaching the lineage-specific expression profile, it becomes more and more clear that epigenetic modifications are highly dynamic. Thus, they might provide an important link between the actions of ROS and cardiovascular diseases. This review will provide an overview of the role of ROS in modulating the epigenetic landscape in the context of the cardiovascular system.
Linked articles: This article is part of a themed section on Redox Biology and Oxidative Stress in Health and Disease. To view the other articles in this section visit http://onlinelibrary.wiley.com/doi/10.1111/bph.v174.12/issuetoc.
© 2017 The British Pharmacological Society.
Figures
References
-
- Ago T, Liu T, Zhai P, Chen W, Li H, Molkentin JD et al. (2008). A redox‐dependent pathway for regulating class II HDACs and cardiac hypertrophy. Cell 133: 978–993. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
