

Parvalbumin Interneurons Modulate Striatal Output and Enhance Performance during Associative Learning
- PMID: 28334608
- PMCID: PMC5386608
- DOI: 10.1016/j.neuron.2017.02.033
Parvalbumin Interneurons Modulate Striatal Output and Enhance Performance during Associative Learning
Erratum in
-
Parvalbumin Interneurons Modulate Striatal Output and Enhance Performance during Associative Learning.Neuron. 2018 Jul 11;99(1):239. doi: 10.1016/j.neuron.2018.06.034. Neuron. 2018. PMID: 30001509 Free PMC article. No abstract available.
Abstract
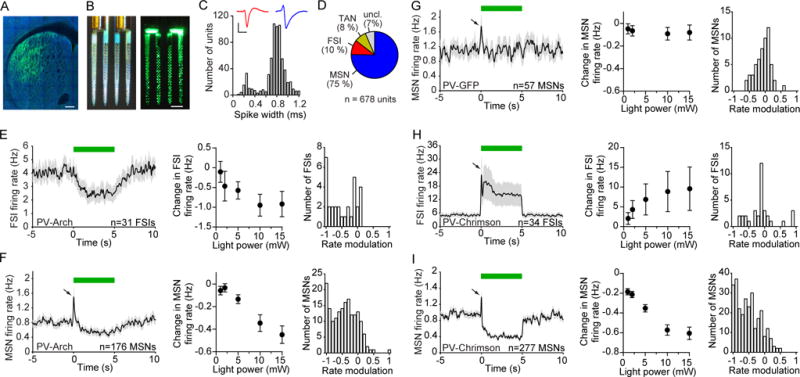
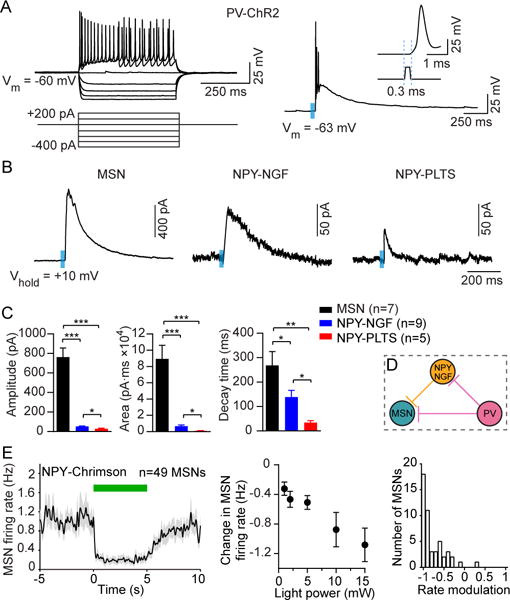
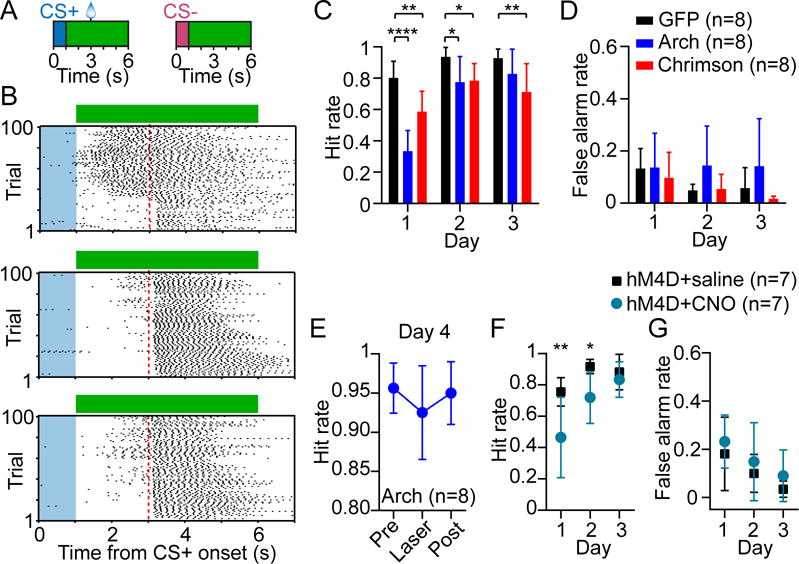
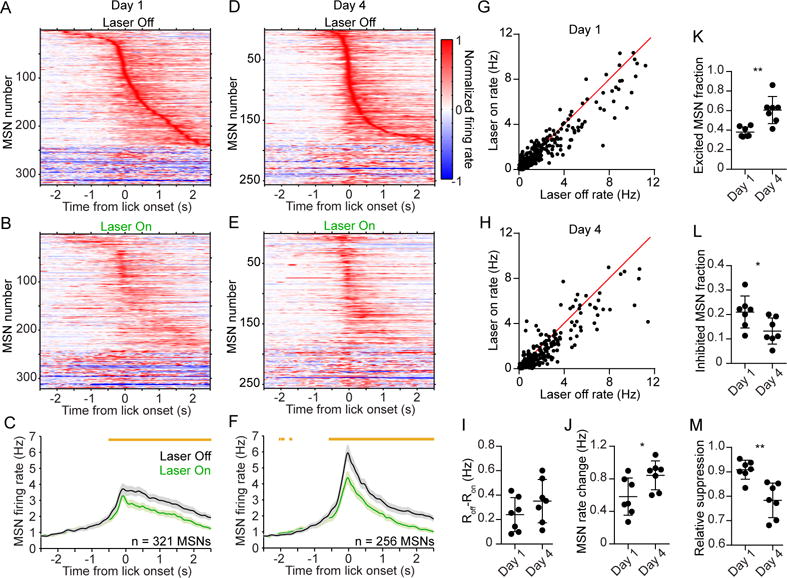
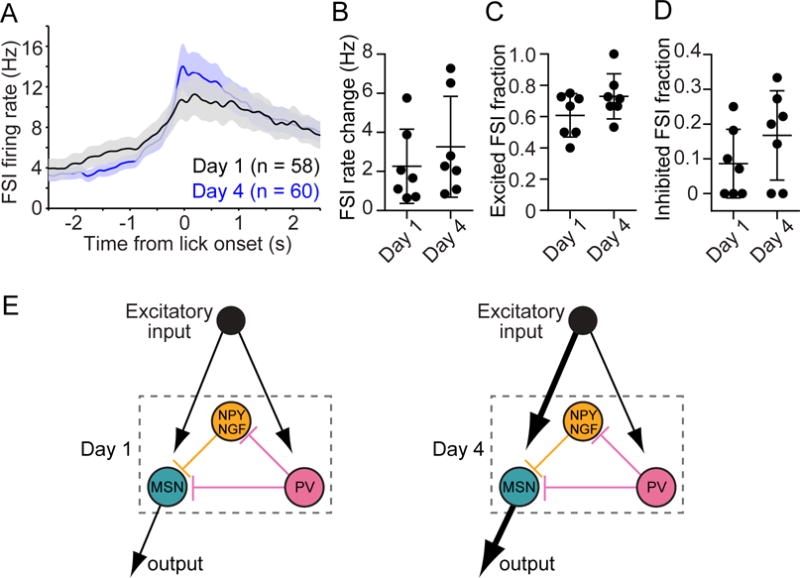
The prevailing view is that striatal parvalbumin (PV)-positive interneurons primarily function to downregulate medium spiny projection neuron (MSN) activity via monosynaptic inhibitory signaling. Here, by combining in vivo neural recordings and optogenetics, we unexpectedly find that both suppressing and over-activating PV cells attenuates spontaneous MSN activity. To account for this, we find that, in addition to monosynaptic coupling, PV-MSN interactions are mediated by a competing disynaptic inhibitory circuit involving a variety of neuropeptide Y-expressing interneurons. Next we use optogenetic and chemogenetic approaches to show that dorsolateral striatal PV interneurons influence the initial expression of reward-conditioned responses but that their contribution to performance declines with experience. Consistent with this, we observe with large-scale recordings in behaving animals that the relative contribution of PV cells on MSN activity diminishes with training. Together, this work provides a possible mechanism by which PV interneurons modulate striatal output and selectively enhance performance early in learning.
Keywords: disynaptic inhibition; learning; neural recording; optogenetics; parvalbumin interneurons; reward conditioning; striatum.
Copyright © 2017 Elsevier Inc. All rights reserved.
Figures
References
-
- Barnes TD, Kubota Y, Hu D, Jin DZ, Graybiel AM. Activity of striatal neurons reflects dynamic encoding and recoding of procedural memories. Nature. 2005;437:1158–1161. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
